ELATERIDARIUM ročník 4 (2010) strany 1-32 (14.01.2010)
Brouci z podčeledi Scolytinae (Coleoptera: Curculionidae) na ovocných dřevinách: literární rešerše
Jana HORÁKOVÁ & Jakub HORÁK
1) Katedra ochrany lesa a myslivosti, Fakulta lesnická a dřevařská, Česká zemědělská univerzita v Praze, Kamýcká 1176, CZ-165 21 Praha, e-mail: xadaj@post.cz 2) Oddělení indikátorů biodiverzity, Výzkumný ústav Silva Taroucy pro krajinu a okrasné zahradnictví, Květnové náměstí 391, CZ-252 43 Průhonice, e-mail: jakub.sruby@seznam.cz
Abstrakt. Cílem této rešerše bylo uceleně zpracovat a utřídit informace o kůrovcovitých z podčeledi Scolytinae troficky vázaných na ovocné dřeviny pěstované v ČR. V našich podmínkách se jedná hlavně o čtyři druhy kůrovců. Dva druhy jsou z rodu Scolytus a patří mezi floeofágní (lýkožravý) hmyz. Jde o bělokaze švestkového (Scolytus mali) a b. ovocného (Scolytus rugulosus). Další dva jsou z rodu Xyleborus a patří mezi xylomycetofágní hmyz – živí se dřevem napadeným houbami. Jde o drtníka ovocného (Xyleborus dispar) a d. všežravého (Xyleborus saxeseni). Všechny tyto druhy napadají dřeviny z čeledi Rosaceae. Drtníci jsou však schopni napadat i jiné listnaté dřeviny. Práce se věnuje jejich bionomii, determinaci dle požerků, areálu výskytu, výživě a vztahu k symbiotickým houbám, jejich predátorům, parazitům ad. A to tak, aby celek popisoval ekologický vztah těchto kůrovců k dřevinám, a zároveň byl použitelný jako ucelený informativní podklad ke studiu těchto kůrovců.
Abstract. The aim of this review was processing and ordinating the knowledge about species of the subfamily Scolytinae feeding on fruit trees in Czech Republic. In Czech Republic dominate four species from two genera. Two species are phloeophagous from genus Scolytus – S. mali and S. rugulosus. Another two species are xylomycetophagous from genus Xyleborus – X. dispar and X. saxeseni. Their most attractive host trees are from family Rosaceae. The woodborers also usually invaded most of the broad leaf tree from other families. The main idea of this paper was to elaborate their ecology – evolution, phenology, number of generations, galleries determination, distribution area, nutrition and symbiotic fungi, predators and parasites etc. Whole should describe their ecological preference to wood plants and should be usable for the study of these beetles.
1.1. Obecný cíl
Cílem této práce bylo uceleně zpracovat a utřídit informace o kůrovcovitých z podčeledi Scolytinae Latreille troficky vázaných na ovocné dřeviny pěstované v České republice (ČR), dále o poškozeních, která způsobují a vytvořit tak rešerši, která by mohla sloužit pro další podrobnější zkoumání druhů.
1.2. Obecné poznatky
1.2.1. Základní charakteristika
Několik druhů kůrovcovitých brouků, vyvíjejících se v lýkové nebo dřevní části ovocných dřevin, může výrazně ovlivnit zdravotní stav ovocných dřevin. Poškození při silnějším napadení může vést k jejich uhynutí.
1.2.2. Výčet hlavních druhů
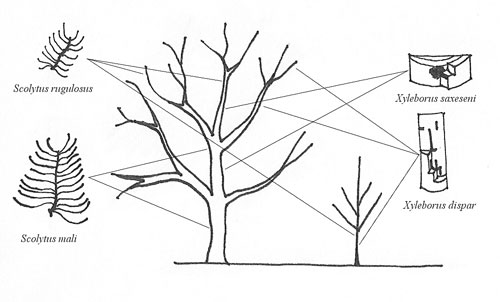
V podmínkách ČR se jedná o čtyři druhy kůrovců. Dva druhy patří mezi floeofágní hmyz a dva mezi xylomycetofágní hmyz. Jedná se konkrétně o bělokaze švestkového (Scolytus mali Bechstein) a b. ovocného (Scolytus rugulosus Müller) z rodu Scolytus Geoffroy, respektive drtníka ovocného (Xyleborus dispar Fabricius) a d. všežravého (Xyleborus saxeseni Ratzeburg) z rodu Xyleborus Eichhoff.
1.2.3. Nejvíce potenciálně ohrožené ovocné dřeviny
Ve většině případů se jedná o druhy z čeledi růžovité (Rosaceae Juss.). Jde o rody hrušeň (Pyrus L.), jabloň (Malus Mill.) a sensu lato rod Prunus L. – slivoň, střemcha, mandloň, mahalebka, bobkovišeň, třešeň, meruňka, broskvoň (syn. Cerasus Mill., Padus Mill., Padellus Vassilcz., Laurocerasus Duh., Amygdalus L., Armeniaca Scop., Persica Mill.) (Kubát 2002). Z dalších rodů připadá v úvahu rod ořešák (Juglans L.) z čeledi ořešákovité (Juglandaceae Kunth.).
2. ROZBOR PROBLEMATIKY
2.1. Systematické rozdělení kůrovců
2.1.1. Poznámky k použitému rozdělení
Kostru v celé práci tvoří rozdělení dle Pfeffera (1989). Je to hlavně z důvodu ucelenosti jeho práce, která je, jako jedna z mála, zaměřena přímo na čeleď kůrovcovití (Scolytidae). Ta však je v dnešním pojetí řazena jako podčeleď čeledi nosatcovití (Curculionidae).
V této části je použito taxonomické rozdělení hlavních druhů kůrovců, kteří mohou působit škody na ovocných dřevinách v podmínkách ČR. V přehledu jsou dále uvedena synonyma názvů (např. Pffefer et Knížek 1993). V literatuře jsou některá užívána a české názvosloví se tak často liší.
Taxonomie podčeledi je zatížena velkou spoustou synonym a homonym na rodové i druhové úrovni, která byla způsobena zvýšeným zájmem entomologů o tuto skupinu především na přelomu 19. a 20. století, kdy kůrovci působili značné škody v lesním hospodářství. Neustálým zpřesňováním názvosloví v souladu s pravidly zoologické nomenklatury zde proto častěji, než v jiných skupinách brouků, dochází ke změnám jmen, mnohdy zažitých a dlouhodobě používaných (Hůrka 2005).
V tomto rozdělení mohou chybět druhy, které jsou v této práci zmiňovány. Vždy jde o druhy, jejichž nálezy na ovocných dřevinách jsou buď sporné (srov. Schwenke 1974), málo významné (Pffefer 1955, Pfeffer 1989) nebo jejich nálezy nepochází z území ČR (Gusev et Rimskij-Korsakov 1953). Všechny tyto druhy jsou zmíněny dále v kapitole 2.5.1.
2.1.3. České a cizojazyčné názvy Scolytus mali – bělokaz švestkový (Lanák et al. 1969, Kutina 1983, Hluchý et al. 1997, Kazda et al. 2003, Hůrka 2005), b. ovocný (Gusev et Rimskij-Korsakov 1953). Nienhaus et al. (1998) uvádí dva názvy b. švestkový a b. jabloňový. Vysoký (1995) uvádí b. švestkový, vypukločnělec švestkový nebo vypuklobřišník švestkový.
SK podkôrnik slivkový/EN large fruit bark beetle/GER grosser Obstbaumsplintkäfer
Scolytus rugulosus – bělokaz ovocný (Lanák et al. 1969, Kutina 1983, Hluchý et al. 1997, Kazda et al. 2003), b. ovocný malý (Gusev et Rimskij-Korsakov 1953).
SK podkôrnik ovocný/EN fruit bark beetle/GER runzliger Obstbaumsplintkäfer
Xyleborus dispar – drtník ovocný (Gusev et Rimskij-Korsakov 1953, Lanák et al. 1969, Novák et al. 1974, Kutina 1983, Voroncov 1986, Vysoký 1995, Hluchý et al. 1997, Kalina 2004, Kazda 2003, Urban 2004, Hůrka 2005).
SK drivnárik ovocný/EN broadleaved pinhole borer/ GER ungleicher Holzbohrer
Xyleborus saxeseni – drtník všežravý (Kutina 1983, Voroncov 1986, Hluchý et al. 1997, Kalina 2004), d. obecný (Gusev et Rimskij-Korsakov 1953). Vysoký (1995) uvádí kromě d. všežravý, též d. hladkoštítý.
SK drivnárik všežravý/EN fruit-tree pinhole borer/ GER kleiner Holzbohrer
2.2. Trofické rozdělení kůrovců
2.2.1. Potravní strategie kůrovcovitých
Řada podkorních druhů hmyzu hostí ve svých požercích bohatá společenstva hub. Nejznámější skupiny se specifickou vazbou na houby patří mezi brouky (Coleoptera). Povaha interakce houby s podkorním hmyzem určuje potravní strategii brouka. V našem mírném pásmu převažuje floeofágie – 90 % druhů kůrovců. V tropech s podmínkami vhodnými pro růst hub je převaha xylomycetofágních (ambróziových) kůrovců – 60 % (Kolařík 2004).
2.2.1.1. Xylomycetofágie
Jedná se o požírání dřevních pletiv (xylemu) napadených houbami (mycelium – podhoubí). Jsou nazýváni též ambróziovými brouky.
2.2.1.2. Floeofágie
Druh konzumuje na rostlině lýkovou část (floem). Šrůtka (2006), zahrnuje obě výše zmíněné strategie pod pojem kambiofágní hmyz.
2.2.1.3. Saprofágie
Živočichové se živí odumřelými a hnijícími zbytky rostlin nebo živočichů. U kůrovců jde zpravidla o saproxylofágii. Což je požírání odumřelého dřeva.
2.2.2. Trofické strategie rodů kůrovců na ovocných dřevinách
U čtyř výše zmíněných kůrovců jde o druhy podkorního hmyzu s určitou vazbou na houby. U druhů z rodu Scolytus jde z hlediska potravní strategie o kůrovce sensu stricto vyznačující se floeofágií. U druhů z rodu Xyleborus jde o ambróziové xylomycetofágní brouky (Beaver 1989).
2.3. Poškození ovocných dřevin
2.3.1. Rozdělení ovocných dřevin
Zřejmě nejlepším ukazatelem z hlediska potencionálního poškození je celkový počet ovocných stromů v ČR. Dále je uveden údaj plochy intenzivních výsadeb v ČR. První údaj není úplně přesný, protože zde pravděpodobně chybí údaje o počtech v extenzivních sadech, které v současné době tvoří jeden z nejlákavějších zdrojů potravy pro kůrovce. Lze předpokládat, že proti níže uvedeným údajům je vyšší zastoupení především u slivoní a ořešáků. Slivoně byly až do 50. let minulého století nejrozšířenější ovocnou dřevinou a jsou stále hojně zastoupeny v dožívajících sadech, často ve formě polykormonů. Ořešáky jsou alejovými stromy ve volné krajině. Naopak nižší zastoupení bude zřejmě u meruněk a broskvoní, které se vyznačují krátkověkostí.
Tabulka 1: Celkový počet stromů a plochy intenzivních ovocných výsadeb v ČR – údaje za rok 1996 (Blažek 1998)
Ovocný druh
% ks
% ha
jabloň
39,6
49,6
hrušeň
4,6
1,8
broskvoň a meruňka
4,6
24,3
slivoně
9,2
1,8
třešně a višně
5,6
14,2
drobné ovoce
34,5
8,0
ořešák
1,8
-
celkem
100 %
(49,8 mil. ks)
100 %
(18 357 ha)
2.3.2. Důsledky napadení kůrovci
2.3.2.1. Stupeň poškození
Druhy kůrovců uvedené v kapitole 2.1.2. nejsou zpravidla v ovocnictví škůdci, kteří by výrazně omezovali produkci. V současné době však byly zaznamenány případy, kdy tyto druhy poškozují dřeviny ve školkách (Urban 2004). Dále mohou škodlivě působit na stromech přeroubovaných na perspektivnější odrůdy, kdy dojde k oslabení stromu. Nejvíce příležitostí stále nalézají v přestárlých ovocných sadech.
Názory autorů na škodlivost kůrovcovitých brouků a jejich larev na ovocných dřevinách nejsou jednotné. Lze konstatovat, že literatura zaměřená na pěstování ovocných dřevin tyto škůdce spíše přehlíží např. Blažek (1998, 2001). Literatura specializovaná na ochranu rostlin se jim věnuje dostatečně. Avšak ve stupni poškození a také v tom, zda jsou škůdci primárními či sekundárními, je nejednotná.
Pfeffer (1955, 1989) uvádí, že některé druhy kůrovců jsou známy jako škůdci dřevin. Při jejich žíru vznikají fyziologická poškození pletiv probíhajících v lýku a běli (viz. 2.4.1.1.) dřevin a napadené dřeviny odumírají. Mezi zástupce potenciálních škůdců v sadech na ovocných stromech patří drtník ovocný (Xyleborus dispar), bělokaz švestkový (Scolytus mali) a bělokaz ovocný (Scolytus rugulosus ).
Většina autorů uvádí tyto kůrovce spíše jako sekundární škůdce, škodící primárně pouze při přemnožení. Kalina (1999) uvádí, že v našich podmínkách jsou to, za normálního stavu, vždy škůdci sekundární – napadající oslabené a odumírající, popř. kompetičně již vyloučené rostliny. Výjimku tvoří pouze druhy poškozující zemědělské plodiny – např. Hylastinus obscurus Marsham, u nás jsou však prakticky bez významu. Z dalších druhů je primární snad již jen rod Thamnurgus. Kolařík (2004) označuje určitou skupinu kůrovců jako primární – např. Dendroctomus micans Kugellanns, Ips typhographus L. nebo Scolytus ratzeburgii Jans. Ti jsou schopni napadnout zdravý strom a zničit ho.
Kutina (1983) uvádí druhy Scolytus mali, Scolytus rugulosus a druh Xyleborus dispar jako velmi závažné škůdce často způsobující velké škody. U jednotlivých druhů uvádí: Scolytus mali – napadá nemocné či mrazem nebo suchem oslabené stromy. Scolytus rugulosus – škodí hlavně na mladých stromcích po výsadbě. Xyleborus dispar – škodí na oslabených ovocných stromech, které pak hynou.
Vážná poškození dřeva mohou způsobit zejména drtníci rodu Xyleborus. Xyleborus dispar je významný především jako technický škůdce, i když někdy napadá stojící fyziologicky oslabené jedince jakéhokoli stáří. Při přemnožení byl pozorován i na zdravých jedincích (Kalina 2004). U druhu Xyleborus dispar uvádí Vysoký (1995), že u nás patří k potencionálním škůdcům v ovocnictví vedle, svilušky ovocné (Panonychus ulmi Koch) a obaleče jablečného (Cydia pomonella L.). U druhu Scolytus mali uvádí pouze, že napadá hlavně stromy nemocné, chřadnoucí po suchých letech, poškozené mrazy, okusem nebo onemocnělé rakovinou. Ohniskem kůrovcové nákazy se mohou stát opuštěné a nepěstované zahrady, odkud se brouci rozletí do okolí. V případě přemnožení napadá i zdravé a mladé štěpy.
Druh Xyleborus dispar se za příznivých klimatických a trofických podmínek může přemnožit a při nedostatku vhodného materiálu napadá i dřeviny zdravé a vitální (Urban 2004). Z dalších autorů např. Hluchý et al. (1997) uvádí, že bělokazové napadají především stromy chřadnoucí v důsledku sucha, mechanického poranění, mrazu, okusu zvěře a podobně. Přičemž silnější napadení může vést až k úplnému uschnutí napadeného stromu. Drtníci napadají především oslabené stromy. Napadení drtníky může vést k usychání celých větví, případně i celých stromů. V případě přemnožení v sadu či vinici napadají drtníci i relativně zdravé stromy či keře révy vinné.
Dřevo a lýko, zvláště starých a oslabených ovocných stromů, poškozují žírem dospělci a larvy kůrovcovitých brouků a to Scolytus rugulosus, Scolytus mali a Xyleborus dispar (Kazda et al. 2003). U drtníků uvádí Lanák et al. (1969), že se vyskytuje především na stromech oslabených jinými škůdci. Škůdce napadá všechny ovocné dřeviny. Tenčí větve usychají nad místy, kde vrtali drtníci. Napadnou-li drtníci silnější větve, následují je kůrovci, jejichž vývoj začíná v místě napadení drtníky, a dále různé houby. Při silnějším napadení strom usychá, slabší napadení strom může přežít, ale je oslaben a živoří. Dále uvádí, že všeobecně převládá názor, že drtníci napadají jen nemocné stromy, popřípadě stromy jinak oslabené, vysazené v nevhodných polohách oslabené nedostatkem živin apod. U bělokaze uvádí, že napadá především stromy nemocné, poškozené mrazem, oslabené suchem apod., a že napadá kmen i silnější větve. Činností tohoto škůdce stromy prosychají a větve během vegetace vadnou. Bělokaz ovocný škodí podobně jako bělokaz švestkový a kromě toho napadá i tenké větvičky v koruně stromu, nesprávně zasazené stromky a hlavně přesazené stromky v mladých výsadbách. Přičemž výskyt bělokazů je dle autorů větší v blízkosti lesa. Rauleder (2003) uvádí, že ovocné plantáže jsou drtníky také ohroženy nejvíce u okrajů lesních porostů. A konečně Urban (2004) uvádí u škodlivosti drtníka ovocného, že jde o široce polyfágního fyziologicko-technického a technického škůdce lesních a zejména ovocných dřevin každého věku. Přednostně napadá churavějící dřeviny na špatných stanovištích, dřeviny oslabené suchem, mrazem, mechanickým poškozením, přesazením, zkrácením kořenů, žírem hlodavců, nevhodným hnojením apod. Je to tedy druh výrazně sekundární, o čemž svědčí jeho častý výskyt v neodkorněných, čerstvě pokácených dřevinách a pařezech. Za příznivých klimatických a trofických podmínek se přemnoží a při nedostatku vhodného rozplozovacího materiálu napadá i dřeviny zdravé a vitální. Osídlené dřeviny často napadají jiní hmyzí škůdci a četné patogenní houby, které pak urychlují jejich zánik. Z ohnisek přemnožení se oplodněné samičky rychle šíří aktivním letem či pasivně (vzdušnými proudy) do blízkého i vzdálenějšího okolí. Hospodářsky závažné škody drtník působí na mladých dřevinách. Silně napadené odrostky nebo dřeviny v růstové fázi mlazin odumírají nebo se v místě poškození lámou pod náporem větru, sněhu, ledovky a námrazy. Z lesních dřevin takto poškozuje hlavně dub, buk, javor a z ovocných dřevin jabloň, hrušeň, švestku a meruňku. Obecně menší význam má technické poškození dřeva. V silnějších kmenech jsou požerky lokalizovány v povrchových vrstvách běle (do hloubky 6 cm) a při pilařském zpracování jsou většinou odstraňovány.
Většina těchto kůrovců je uváděna autory jako čistě sekundární škůdci. Xyleborus saxeseni při dostatku vlhkosti ve dřevě může silně napadnout odkorněné dřevo (Kalina 2004). Scolytus mali se vyskytuje i v pařezech a pokácených nebo polámaných kmenech (Vysoký 1995). Xyleborus dispar nalétává v květnu na čerstvé pařezy, na odumírající stromy a na pokácené neodkorněné dříví.
2.4. Ekologická nika jednotlivých druhů
2.4.1. Nika jednotlivých rodů
2.4.1.1. Rozdělení dřeva (xylemu)
Dřevo v živém stromě má dvojí úlohu. Fyziologickou, která je spojena s rozvodem vody a živin z kořenů do dřeviny. Tato úloha náleží mladším obvodovým částem dřeva – běli. Ta se liší zpravidla světlejší barvou od tmavších částí – jádra. Buňky jádrového dřeva jsou vesměs odumřelé a plní mechanickou úlohu. Význačně barevné jádro má většina ovocných dřevin, například ořešák, švestka, třešeň nebo jabloň (Kofránek 1950).
2.4.1.2. Rozmístění rodů v dřevině
Oba druhy rodu Scolytus patří mezi fyziologické škůdce. Jejich požerek probíhá pod kůrou v lýku a je částečně zaříznut do běle, přičemž hloubka v běli koreluje s vlhkostí dřeva (Lanák et al. 1969). Kukelné kolébky zasahují hlouběji do běle (Gusev et Rimskij-Korsakov 1953, Lanák et al. 1969, Vysoký 1995).
Oba druhy rodu Xyleborus patří mezi fyziologicko-technické škůdce. Jejich požerek probíhá v dřevní části, většinou je to běl, výjimečně zasahují chodby do jádra.
2.4.2. Nika jednotlivých druhů
U všech čtyř druhů kůrovců je z obecného hlediska víceméně podobně rozdělena nika. U Scolytus rugulosus a Xyleborus dispar je to tenčí dřevo. U Scolytus mali a Xyleborus saxeseni je to tlustší dřevo. I zde jsou autoři často nejednotní. U druhu Scolytus mali někteří uvádí, že se vyskytuje i na tenkém dřevě. Je však nutné poznamenat, že se jedná zpravidla o literaturu obecnější (Hůrka 2005, Kazda et al. 2003). U rodu Xyleborus jsou názory ještě nejednotnější a proto výše zmíněné rozdělení je spíše obecné a platí hlavně za předpokladu, že se jedná o ovocný strom, který je obsazen oběma druhy drtníků (Horák 2007a).
Preference kůrovců k dimenzím dřeviny a jejich požerky.
2.4.2.1. Druhy rodu Scolytus
Gusev et Rimskij-Korsakov (1953) uvádějí, že u druhu Scolytus mali jsou larvové chodby podél kmene v tlusté kůře. Kutina (1983) uvádí u tohoto bělokaza žír na kmenech a tlustých větvích pod kůrou. Naopak u Scolytus rugulosus uvádí, že škodí hlavně na mladých stromcích. Gusev et Rimskij-Korsakov (1953) u něj uvádí žír na větvích a kmenech s tenkou kůrou. Lanák et al. (1969) uvádějí, že bělokaz ovocný škodí podobně jako bělokaz švestkový a kromě toho napadá i tenké větvičky v koruně stromu, nesprávně zasazené stromky a hlavně přesazené stromky v mladých výsadbách. A konečně Vysoký (1995) uvádí, že Scolytus mali se vyvíjí pod kůrou dřevin, nejčastěji kmenů a silných větví, často s Scolytus rugulosus s nímž má shodný vývoj, ale Scolytus rugulosus je ve slabších větvích a kmíncích.
2.4.2.2. Druhy rodu Xyleborus Xyleborus dispar se vyvíjí v čerstvě vlhkých vývratech, zlomech a pařezech všech dimenzí. V tenkých kmíncích a větvích matečné chodby často vybíhají z krátké vstupní chodby, a to v různé celkové délky (obvykle 2 až 4 cm). V silnějších kmenech jsou požerky lokalizovány v povrchových vrstvách běle (do hloubky 6 cm) (Urban 2004). Hůrka (2005) uvádí, že druh se vyvíjí ve větvích i kmenech téměř všech listnatých stromů. Naopak Kutina (1983) uvádí, že Xyleborus dispar se vyvíjí na 2–3letém dřevě, kde jsou jednotlivé kruhovité otvory. Novák et al. (1974) uvádí, že letní závrty na mladých listnáčích, popř. na větvích starších stromů se připisují rojení a opakované snůšce rodičovských brouků, kteří opouštějí kolonie od doby, kdy se v nich objeví první kukly a mladí brouci.
U druhu Xyleborus saxeseni uvádí Kutina (1983), že chodba směřuje radiálně do středu větve a rozšiřuje se. Jeho vstupní chodby o průměru kolem 1 mm pronikají kolmo do dřeva a do hloubky 1 až 3 cm. Na rozdíl od ostatních drtníků se jeho larvy živí zčásti i dřevem, neboť hlavně v měkkém jarním dřevě vyžírají ploché, kapsovité dutiny (Urban 2004).
2.5. Další druhy podkorního hmyzu
I přesto, že většinu ovocných dřevin, lze jen stěží označit za původní dřeviny, z entomologického hlediska jsou hostiteli celé řady hmyzích druhů a jejich vývojových stádií. Gusev et Rimskij-Korsakov (1953) uvádí na ovocných dřevinách celkem devět druhů jen z podčeledi Scolytinae. Schwenke (1974) jich uvádí nejméně osm. V našich podmínkách pravděpodobně připadá v úvahu pět až šest druhů (Pfeffer 1989) a z toho čtyři jsou běžní.
2.5.1. Další kůrovcovití
Z dalších druhů kůrovců připadá v úvahu na lísce (Corylus avellana L.) Scolytus carpini Ratz. (Gusev et Rimskij-Korsakov 1953, Schwenke 1974). Dále Scolytus amygdali Guér., který žije na meruňce (Prunus armeniaca L.), a jehož podélné matečné chodby jsou otisknuty jen na vnitřní straně kůry, larvové chodby jsou řídké a procházejí zpočátku na spodní straně kůry, později vnikají do její horní vrstvy, kde jsou rovněž kukelné kolébky (Gusev et Rimskij-Korsakov 1953). Na hrušních (Pyrus) a rodu Prunus uvádí Pfeffer (1989) druh Scolytus laevis Chapuis, který má pokolení jednoleté a v pahorkatinách je dosti hojný. Schwenke (1974) ho uvádí pouze na rodu Prunus. Na třešních (Prunus avium L.) a višních (Prunus cerasus L.) se vyskytuje nepříliš hojný druh kůrovce Polygraphus grandiclava Thomson, jde o polygamní druh, jehož dalšími živnými dřevinami jsou některé jehličnany (Pfeffer 1989). Poměrně specifickou dřevinou je z hlediska podkorních škůdců ořešák vlašský (Juglans regia L.). Gusev et Rimskij-Korsakov (1953) na něm uvádí další tři druhy, konkrétně druh z tribu Xyloterini Lindemann Xyloterus domesticus L, který má žebříčkovité požerky. Jeho matečné chodby jsou oproti druhu Xyleborus dispar delší a probíhají většinou v příčné rovině. Ve směru dřevních vláken z nich odbočují nahoru a dolů kratičké larvové chodbičky o délce kolem 5 mm. Dřevokaz bukový poškozuje hlavně buk a jiné tzv. tvrdé listnáče, velmi zřídka ovocné stromy (Urban 2004). Dále autoři uvádějí druhy Scolytus scolytus Fabr., který je znám jako vektor tracheomykózy u jilmů a dále z podčeledi Hylesininae Erichson druh Leperisinus fraxini Panzer jehož matečné chodby pod kůrou jsou příčné. Druh Scolytus scolytus Fabr. uvádí Schwenke (1974) i na rodu Prunus na kterém uvádí ještě dva druhy bělokazů Scolytus multistriatus Marsh. a Scolytus kirschii Skal. Posledním zástupcem je Xyleborus monographus F., kterého Schwenke (1974) uvádí na druzích Juglans regia L., Prunus avium L. a Castanea sativa Mill. Urban (2004) ho však na ovocných dřevinách vylučuje.
2.5.2. Další druhy hmyzu a symptomy jejich poškození
Z dalších druhů hmyzu lze jmenovat druhy, které působí významnější škody, jako jsou zástupci řádu Lepidoptera. Například dva zástupci drvopleňů – drvopleň obecný (Cossus cossus L.) a d. hrušňový (Zeuzera pyrina L.). První jmenovaný škodí hlavně na jabloních, hrušních, slivoních a ořešáku. Kromě nich se vyskytuje na vrbách a topolech. Jde o velký druh nočního motýla, jehož rudožluté housenky dosahují i přes 10 cm. Vyhlodávají nepravidelné chodby, ze kterých vyhazují trus. Napadené dřevo zapáchá po octě. Napadá hlavně staré stromy, ale zaznamenán byl i v souvislosti s používáním gumových chráničů proti okusu zvěří. Kuklí se zpravidla přímo v napadeném dřevě. Druhý jmenovaný druh napadá hlavně mladší stromy. Zpravidla jde o hrušně, jabloně nebo ořešáky. Nejčastěji se však vyskytuje v jasanech. Prvý instar žere v listech a letorostech, pak proniká pod kůru, kde hlodá plošné chodby přecházející postupně do dřeva. Kukla vyčnívá z otvoru ve dřevě. Žír housenek prozrazují třísky promíšené s kuličkovitým trusem vytlačované z vyvrhovacího otvoru. Dospělí noční motýli jsou pěkně bíločerně vybarvení (Hluchý et al. 1997, Kalina 2004). U obou druhů jde o xylofágní hmyz, jehož vývoj je zřejmě vzhledem k nízké nutriční hodnotě dřeva víceletý.
Dalším zástupcem motýlů je obaleč meruňkový (Enarmonia formosana Scopoli), který je jedním z původců apoplexie (mrtvice) u meruněk. Jeho housenky vyžírají pod kůrou nepravidelné chodbičky, z poškozených míst strom roní mízu a hojením ran vznikají nádorovité novotvary (Kazda et al. 2003).
Již méně významní jsou nesytkovití (Sesiidae). Tito motýli připomínající blanokřídlý hmyz jsou převážně ranovými škůdci. Jejich žír probíhá pod kůrou, je zpravidla víceletý a po vylíhnutí lze povětšinou u výletového otvoru nalézt exuvii kukly (Hluchý et al. 1997, Kalina 2004). Větší škody působí hlavně na drobném ovoci (Horák 2007b).
Další skupinou je hmyz, který se živí na ovocných dřevinách, ale je spíše zajímavý a vzácný (Horák 2007d). Z krascovitých (Buprestidae) jsou to například překrásně vybarvení – krasec třešňový (Anthaxia candens Panzer), krasec (Anthaxia fulgurans Schrank) nebo krasec lesknavý (Anthaxia nitidula L.) jejichž larvy se vyvíjí pod kůrou odumřelých osluněných částí ovocných stromů. Dalším zástupcem je u nás již vyhynulý Capnodis tenebrionis L., jehož larvy mají dlouhý vývoj probíhající v kořenech dřevin z rodu Prunus L. Z tesaříkovitých (Cerambycidae) je to například dříve poměrně běžný tesařík bukový (Cerambyx scopolii Füessly), s až 5 cm velkou larvou žijící ve dřevě zejména třešní nebo atraktivně zbarvený tesařík broskvoňový (Purpuricenus kaehleri L.), který je u nás z 3 zástupců rodu nejrozšířenější, přesto však vzácný. Jde o teplomilný druh, jehož larvy se vyvíjí v tenkých větvích především hrušní, broskvoní a meruněk (Hůrka 2005).
2.5. Doporučené prameny
Pro tuto rešeršní práci jsme se snažili využít především literaturu domácí. Pro případné objasnění některých tvrzení či pro rozšíření znalostí týkajících se kůrovcovitých vázaných na ovocné dřeviny lze doporučit ještě další domácí prameny, jako jsou práce Pfeffera (1942, 1954). Další informace se nachází i v cizojazyčných pracech ať již starší (Karpiňski et Strawiňski 1948, Balachowsky 1949, Stark 1952, Nunberg 1954), ale i novější (Wood 1982, Pfeffer 1995, Kolk et Starzyk 1996, Dominik et Starzyk 2004).
3. MATERIÁLY
3.1 Kůrovcovití (Scolytinae)
V současné době jsou kůrovcovití řazeni do čeledi nosatcovití (Curculionidae). Ta se tím stala druhově nejbohatší čeledí brouků, ale zároveň i pro laika téměř nepřehlednou (Knížek 2008). Proto je zde použit klíč, který je založen právě na již zmiňovaném díle Pfefferově (Pfeffer 1989), kde jsou kůrovcovití řazeni jako samostatná čeleď.
3.1.1 Klíč nadčeledi Scolytoidaea Pearce
1 (2) Prvý článek chodidel delší než ostatní chodidlové články do¬hromady, a tak dlouhý jako holeň. Hlava shora viditelná a tak široká jako přední okraj štítu. Štít dlouze válcovitý a po stra¬nách předního okraje rovně dolů zkosený. Tělo dlouze válcovi¬té.
…čeleď Platypodidae Latreille
2 (1) Prvý článek chodidel kratší než druhý a třetí článek dohroma¬dy, kratší než holeň. Hlava užší než přední okraj štítu. Shora je viditelná z části, nebo zcela neviditelná. Štít po stranách před¬ního okraje šikmo dolů zkosený. Tělo oválně zaoblené, nebo krátce válcovité.
…čeleď Scolytidae Latreille
3.1.2 Klíč čeledi Scolytidae Latreille
1(4) Přední tibie na vnější straně ozubené, nezakončené na vnější straně trnem. Krovky silně klenuté, kryjící po stranách zadohruď a zadeček.
…2
2(3) Báze krovek poněkud vyzdvižená, a v těchto místech s hrbolky nebo zoubky. Je-li okraj krovek výjimečně hladký (Hylastini LeConte), pak je hlava protažena v krátký nosec. Štít vždy tečkovaný. Konec krovek zaoblený, bez zoubků. Na jehličnanech, listnáčích i bylinách.
…podčeled Hylesininae Erichson
3(4) Báze krovek není vyvýšená. Štít v přední části s drobnými hrbolky, v zadní části tečkovaný, jen výjimečně vpředu i vzadu tečkovaný (Crypturgini LeConte a Thamnurgini Nüsslin). Na jehličnanech i listnáčích.
…podčeled Ipinae Reitter
4(1) Vnější strana předních tibií hladká, konec tibií s vyčnívající ostruhou. Krovky ploché, strany zadečku dobře viditelné, poslední články ubíhající strmě šikmo vzhůru. Štít tečkovaný. U nás výlučně na listnáčích.
…podčeled Scolytinae Latreille
3.1.3. Biologie
Všichni zástupci této čeledi se vyznačují určitou péčí o své potomstvo. Dospělci vyhledávají při zakládání nového pokolení ochořelé nebo odumírající dřeviny, případně byliny. U monogamických druhů se zavrtává do kůry napřed samička a vylučováním feromonu přiláká samečka. Zástupci rodu Scolytus kopulují na povrchu kmene. U druhů polygamních se naopak do kůry zavrtává nejprve sameček. V kůře nebo v lýku vyhlodává rozšířenou snubní komůrku a vylučovaným feromonem přiláká samičky. Většina druhů kůrovců po obsazení dřeviny vylučuje do ovzduší agregační feromon, kterým signalizuje vhodné prostředí i pro další jedince téhož druhu, takže následuje hromadný nálet kůrovců na dřevinu. Ojedinělým náletem se vyznačuje lýkohub Dendroctomus micans Kug.
Samičky poté hlodají v lýku, nebo i v běli tzv. matečné chodby, kde kladou postupně svá vajíčka a chodby postupně prodlužují. Vylíhlé larvy hlodají pak v lýku do stran své vlastní larvové chodby. Dorostlé larvy rozšiřují konec chodby v kolébku, kde se kuklí. Larvy se při svém žíru živí buď pouze částí pletiv živných rostlin, nebo se přikrmují podhoubím některých druhů hub, které zde bují nebo se vyživují pouze podhoubím ambróziových hub, které porůstá stěny chodeb vyhlodaných v běli stromů. Spory těchto hub přenášejí samičky ve zvláštním orgánu (mycangiu) z míst, kde se vylíhly.
Matečné a larvální chodby kůrovců tvoří pod kůrou nebo ve dřevě výrazně utvářené požerky, které jsou charakteristické pro jednotlivé druhy této čeledi. Lze je rozlišit v požerky jednoramenné svislé, jednoramenné vodorovné, víceramenné svislé, svorkovité nebo hvězdicovité a v požerky plošné. Závrty mateřských brouků se nalézají obvykle v prasklinách kůry nebo pod šupinkami borky. Kolem těchto závrtů se hromadí hnědavě zbarvené drtinky kůry, u dřevokazných druhů bělavé, práškovité drtinky dřeva. U zástupců rodu Tomicus Latr. a Dendroctonus Er. se kolem závrtů objevují bělavé krupičky pryskyřice smíšené s drtinkami kůry.
Zástupci rodu Xyleborus Eichh. a Xyloterus Er. vyhlodávají své matečné chodby v běli stromů. Chodby jsou trubicovité jednoduché nebo se větevnatě dělí v rovině kolmé na osu kmene a někdy i v nejrůznějších směrech. Zřídka je matečná chodba svisle dlanitě rozšířená. Dřevokazové mají matečné chodby utvářené žebříčkovitě, z vodorovně probíhající matečné chodby odbočují nahoru i dolů kratičké chodby larvové. Larvy všech dřevokazných kůrovců se neživí dřevem, ale podhoubím ambróziové houby Leptographium lundbergii Webb. (podle Šrůtky (2006) jde o druh Ambrosiella hartigii Batra), které porůstá stěny chodbiček, dokud je dřevo dostatečně vlhké. Spory houby přenášejí samičky v mycangiích ve štítu a umisťují je na stěny čerstvě vyhlodaných chodeb.
Mycetangia (=mycangia) jsou dutinky v kutikule dospělců, ve kterých jsou přítomné funkční žláznaté buňky, v nichž se přenáší ambróziové stadium houby. Sekrety těchto žláz podporují růst ambróziových hub a potlačují ostatní druhy. Nepřítomnost mycetangií u některých ambróziových brouků ukazuje, že nejsou pro šíření hub nezbytná a jejich tlustostěnné spory mohou být šířeny i v zažívacím traktu (Kolařík 2004).
Hůrka (2005) uvádí, že žijí v jehličnatých a listnatých stromech nebo keřích jen několik druhů se vyvíjí v bylinách nebo semenech rostlin. Dorozumívání jedinců je založeno na chemické komunikaci pomocí feromonů. Kůrovci mají rozvinutou péči o potomstvo. Živí se rostlinnými tkáněmi, někteří ambróziovými houbami, které rostou na stěnách larválních chodeb. Při přemnožení se některé druhy mohou stát vážnými škůdci dřevin a kulturních rostlin.
Voroncov (1986) uvádí, že každý druh kůrovce se vždy usídluje na určitém druhu dřeviny nebo na několika příbuzných druzích a obsazuje určitou část stromu. Rojení kůrovců obvykle začíná zjara a pak trvá do poloviny léta. Nejdříve (konec IV) začínají létat lýkohubi, pak kůrovci a nakonec bělokazi, kteří žijí na listnatých dřevinách.
Všichni kůrovci zakládají v období rozmnožování rodinu. Přitom některé druhy kůrovců (lýkohubi a bělokazi) tvoří monogamní rodiny, složené ze samičky a samečka a jiné (lýkožrouti) tvoří polygamní rodiny, složené z jednoho samečka a několika samiček. Zvláštnosti kůrovcovitých rodin těsně souvisí se zmíněnými rozdíly ve stavbě jejich těla.
Ze snubní komůrky může vést několik matečných chodeb, jejichž počet je pro každý druh charakteristický. Vyskytují se i druhy, které vyhlodají závrtový otvor, končící krátkou matečnou chodbou, v níž samička klade vajíčka na hromádky (Dendoctomus micans Kug., Xyleborus saxeseni aj.). Nejmenší kůrovci rodu Crypturgus osidlují chodby jiných kůrovců a z jejich krajů začínají budovat své vlastní požerky tvářící hustou síť.
Rojení a kladení trvá asi měsíc. Stádium vajíčka trvá asi 10–14 dnů, stadium larvy 15–20 dnů, stadium kukly 10–14 dnů. To znamená, že vývoj je ukončen za 1,5–2 měsíce. Pak začíná období dodatečné výživy, které je nezbytné pro dokončení rozvoje pohlavní soustavy. U většiny kůrovců probíhá pod kůrou stromu, kde vykusují krátké chodbičky různého tvaru. Některé druhy vykusují vnitřní část mladých výhonků. Ty se pak lámou a padají na zem. Mnoho bělokazů se živí šťavnatým lýkem ve vidlicích větviček, někteří obyvatelé kořenů ohryzávají kmínky mladých borovic.
Většina kůrovců má během roku jedno nebo dvě pokolení. Stává se, že larvičky druhé generace se nestačí přeměnit na brouky, přezimují pod kůrou a při nižší teplotě zmrznou. Odolnost kůrovců proti nízkým teplotám těsně souvisí se způsobem zimování. Kůrovci, kteří mají jedno pokolení do roka, zimují ve stádiu brouka u báze stromů v krátkých komůrkovitých chodbičkách, v kůře nebo hrabance. I když nastanou nízké teploty, jsou pod sněhovou pokrývkou málo zranitelní. Mladí brouci, kukly a larvy, zimující pod kůrou stromů ve svých požercích, snášejí nízké teploty různě. Jestliže je takové zimování u druhu obvyklé, larvy snášejí teplotu až –30 °C, jestliže takové zimování není obvyklé, pak mnohé larvy hynou již při –15 °C. Příčinou úmrtnosti larev a kukel mohou být i vysoké letní teploty, obvykle nad horní teplotní hranici vývoje kůrovců 40 °C. Další možnou příčinou úhynu přezimujících stádií jsou dlouhá a vlhká jarní období, kdy dochází k zaplesnivění brouků.
Vysoký (1995) navíc uvádí, že larvy se vyvíjejí ve kmenech a ve větvích různých druhů dřevin i ve stoncích některých rostlin. Při přemnožení se stávají nebezpečnými škůdci lesních i ovocných dřevin. Kůrovci létají pouze v době kladení vajíček, a to vždy za jasného a slunného počasí. Bíle zbarvená vajíčka mají jemný obal. Stádium vajíčka trvá 10–14 dní. Mladí brouci čerstvě vylíhlí z kukel, jsou nevýrazně zbarveni a nemají úplně vyvinutou pohlavní soustavu a k nabytí pohlavní zralosti potřebují zralostní žír. Svým způsobem zralostního žíru často velmi zvyšují svou škodlivost. Přezimují jako imago, kukla nebo larva.
Kalina (1999) uvádí, že rojení dospělců je hromadný jev. Jarní rojení probíhá zpravidla v odpoledních hodinách, letní rojení je nejvýraznější těsně kolem západu slunce. Bělokazové jsou však vždy nejčilejší v nejteplejších denních hodinách. Následuje vyhledání vhodné rostliny – stromy a keře oslabené, churavějící, pokácené a neodkorněné, polámané, jindy zdánlivě zdravé (Dendroctonus – stromy s červenou hnilobou) nebo byliny zcela zdravé (Thamnurgus). Zatímco v první fázi náletu rozlišují kůrovci spektrum silic vylučovaných stromem, v další části převládá orientace podle agregačních feromonů. Rozmnožují se oplozenými vajíčky. Ve vybraném stromě začínají mateřští jedinci hlodat mateřskou komůrku. Které pohlaví tuto činnost zahajuje, závisí na druhu kůrovce. U druhů monogamních a u rodu Xyleborus začíná samička, u polygamních samečkové. Mnoho jich je zalito pryskyřicí, proto se často mění poměr pohlaví ve prospěch samiček, brouci proto využívají symbiotických hub z rodu Ophiostoma a Ceratocystis, které dovedou zablokovat tok pryskyřic (Farrel et al. 2001). Po vyhlodání snubní komůrky u polygamních kůrovců přilétají za samečkem samičky a dochází v komůrce k jejich oplodnění. Poté samičky vyhlodávají mateřské chodby ze snubní komůrky. Směr těchto chodeb je poměrně striktně dodržován, takže vznikají požerky druhově dosti charakteristického typu. Prodloužení chodby, couvání do snubní komůrky, obrat, couvání, kladení vajíčka a znovu. U druhů ve dřevě jsou vajíčka kladena jen na obvod chodby (drtníci rodu Xyleborus) nebo do zářezů (dřevokazové rodu Xyloterus), materiál je infikován ambrosiovými houbami. Jinde jsou kladena vajíčka do mělkých zářezů velké snubní komůrky ve skupinách – Cryphalus Er., Dendroctonus Er., jinde ve skupinách na různých místech mateřské chodby (Hylurgops glabratus Zetterstedt). Pravidelnost a rozestupy, stejně jako orientace mateřské chodby jsou charakteristickými druhovými znaky. Delší mateřské chodby jsou opatřeny dýchacími otvory. U monogamních druhů vyhlodává začátek chodby s rozšířením (snubní komůrku) samička, za ní přilétá sameček a dochází k oplodnění. Druhy vyvíjející se ve dřevě nemají žádnou snubní komůrku. Samičky jsou zde oplodňovány u závrtu (Xyloterus Erichson) nebo přímo v místě, kde se vylíhly (Xyleborus). Samečkové hynou brzy po kopulaci, samičky žijí daleko déle v souvislosti s potřebou vyklást vajíčka. Mnoho druhů kůrovců má pouze jednu generaci do roka (např. Hylastes Erichson, Hylurgops LeConte, Xyloterus Erichson), mnohé však i 2–3 generace do roka (druhy rodu Ips DeG., Pityogenes Bed., Polygraphus Er. a další) a spíše výjimečně je vývoj dvouletý (např. Dendroctonus micans Kugelanns, Hylesinus crenatus F.) (Kalina 1999).
3.1.4. Hospodářský význam
Některé druhy kůrovců jsou známy jako škůdci dřevin a kulturních rostlin (Švestka et al. 1998). Při jejich žíru vznikají fyziologická poškození pletiv a napadené dřeviny i byliny pak rychle odumírají. Mnohé druhy kůrovců se přemnoží při polomech způsobených větrem, sněhem nebo námrazou a pak napadají i zdravé stromy v okolních porostech. Při jejich žíru vznikají fyziologická poškození pletiv probíhajících v lýku a běli dřevin, a napadené dřeviny odumírají. V sadech na ovocných stromech škodí drtník ovocný (Xyleborus dispar), bělokaz švestkový (Scolytus mali) a bělokaz ovocný (Scolytus rugulosus). Kůrovci poškozují pokácené dřevo v lese, ve skladech, ale i technicky. Při přemnožení poškozují takto i oslabené stromy v lesních porostech. Na dubech takto škodí hlodáním chodeb ve dřevě drtník ovocný (Xyleborus dispar), drtník prostřední (Xyleborus monographus F.), drtník dubový (Xyleborus dryographus Ratzb.), drtník všežravý (Xyleborus saxeseni), dřevokaz dubový (Xyloterus signatus F.) a dřevokaz bukový (Xyloterus domesticus L.). Na ostatních listnáčích nejčastěji dřevokaz bukový (Xyloterus domesticus L.), drtník všežravý (Xyleborus saxeseni) a drtník ovocný (Xyleborus dispar).
Některé druhy kůrovců jsou známy i jako přenašeči spor těch druhů hub, které působí hromadné odumírání dřevin. Mezi přenašeče původce tracheomykózy (grafiózy) všech našich druhů jilmů patří bělokaz pruhovaný (Scolytus multistriatus Marsh.), bělokaz jilmový (Scolytus scolytus F.), bělokaz malý (Scolytus pygmaeus F.), bělokaz chlumní (Scolytus laevis Chap.), bělokaz (Scolytus ensifer Eichh.), lýkohub jilmový (Pteleobius vittatus F.) a lýkohub tečkovaný (Pteleobius kraatzi Eichh.). Bělokaz dubový (Scolytus intricatus Ratzb.) přenáší spory hub, které zřejmě vyvolávají tracheomykózní onemocnění dubu zimního a d. letního.
Současné poznatky však hovoří o tom, že vývoj tohoto kůrovce probíhá na již odumřelém materiálu a pouze úživným žírem se podílí na poškození (Zahradník 2006).
Vysoký (1995) uvádí, že hmyz technicky škodí zpravidla v larválním stádiu a jen ojediněle také jako dospělci (např. imaga některých kůrovců). Výskyt dřevokazného hmyzu zjistíme zpravidla až po jeho vylétnutí z napadeného dřeva, protože se objeví na povrchu jeho výletový otvor. Podle tohoto otvoru se dá již orientačně odhadnout, o jaký druh hmyzu se jedná. Pro přesné určení je však většinou nutno mít k dispozici imago.
3.1.5.1. Hmyzí kalamity, kontrola a prognóza
Na rozmnožování hmyzu mají vliv jak činitelé vnitřní, vlastní určitému druhu hmyzu, tak vnější, vyplývající z prostředí, v němž daný druh žije. Ti tak jednak jeho rozmnožování podporují, jednak brzdí, čehož důsledkem je určitý vyrovnaný stav. Dojde-li k změně některých podmínek, které se začnou utvářet příznivěji ve prospěch hmyzu, mívá to za následek stupňování jeho početního stavu – gradaci. Ovšem v přírodě se vyskytnou zřídka optimální podmínky. Je-li jeden činitel v optimu, ostatní, činitelé naproti tomu bývají méně nebo více od optima vzdáleni.
Neškodný stav určitého škodlivého hmyzu je nazýván železnou zásobou. Vzestup počtu jedinců od železné zásoby až po vyvrcholení stavu a dále pokles až na nejnižší stav je gradace hmyzu. Během gradace rozeznáváme několik údobí: stav latence, za kterého počet jedinců po přechodnou dobu je nižší než železná zásoba, dále období, během něhož pozvolným stoupáním dosáhne stav železné zásoby – tj. akrescence, po ní následuje rychlejší vzestup, tj. progrese, kdy dojde k překročení železné zásoby, a vrcholí v kulminaci, kdy počet jedinců dosahuje maxima. Tento stav kulminace netrvá dlouho, je vystřídán rychlým poklesem až k hranici železné zásoby – údobím regrese, kdy ještě pak pozvolna klesá tento stav pod železnou zásobu, to je údobí dekrescence, vystřídané nejnižším stavem jedinců, údobím latence. V životě hmyzu je možno pozorovat stále tyto výkyvy, střídání minim, normálních stavů, přemnožení a opět zase poklesu. Přemnožením hmyzu nastává epidemie a vážná epidemie je nazvána kalamitou. V údobích mezi kalamitami jsou léta normální, tj. stav latence, léta inkubace, tj. nápadný vzestup nad hranici železné zásoby, léta erupční tj. vrcholení epidemie a léta krize, tj. pokles až po hranici železné zásoby. O vzniku hmyzích kalamit bylo vypracováno několik hypotéz, které vysvětlují příčiny přemnožení škůdců (Pffefer 1961, Vyskot 1962).
3.1.6. Zastoupení ve fauně
Řád brouci (Coleoptera – kolem 300 000 druhů), do kterého kůrovcovití náleží, je druhově nejbohatší řád hmyzu (ca 6 500 druhů v ČR, v ca 86 čeledích). Patří do něj kolem 40 % všeho známého hmyzu. Zahrnuje tak asi 1/3 celkového počtu všech známých živočišných druhů (Kalina 1999).
Z více než 6 000 známých druhů kůrovců je jich z ČR a SR uváděno 120 (Hůrka 2005). Pfeffer et Knížek (1993 in Vysoký 1995) uvádí, že na celém světě je známo více než 5 700 druhů kůrovců, z nich připadá na tropy asi 75% a na Evropu kolem 5%. V Palaearktu je známo na 60 rodů a kolem 250 druhů. V ČR je v současnosti známo 104 druhů a jeden druh adventivní. Pro zajímavost pro tehdejší SSSR uvádí Voroncov (1986) 300 druhů kůrovců.
3.1.7. Sběr a odchyt kůrovců
Sběr kůrovců se liší od běžných způsobů sběru jiného hmyzu. Dospělci kůrovců tráví většinu svého života v pletivech živných dřevin a bylin. Brouky, kteří zakládají nové pokolení, nacházíme ve vyhlodaných matečných chodbách v lýku a v běli živných dřevin. Nově vylíhlí jedinci většiny druhů kůrovců hlodají pak ještě v místech, kde se vylíhli svůj zralostní žír, který trvá i několik měsíců. Pouze zástupci rodu Scolytus Geoffr., Hylesinus F., Leperisinus Reitt., Pteleobius Bed. a Tomicus Latr. opouštějí záhy po vylíhnutí požerky a zalétají na živé stromy, kde hlodají svůj zralostní žír ve výhoncích anebo v kůře při bázi kmenů. Venku nacházíme kůrovce pouze po krátký čas při jejich rojení, kdy nalétávají na kůru vyhlédnutých stromů. Proto lze nejlépe vyhledávat kůrovce v živných dřevinách a bylinách během celého roku. K tomu je ovšem třeba znát nejen jejich živné dřeviny a byliny, ale i orgány dřevin, kde se vyvíjejí a také i stupeň zavadání živné rostliny (Pfeffer 1989).
3.1.7.1. Zjišťování stavů kůrovců
Lapáky jsou jedním z nejužívanějších prostředků kontroly, jimiž kontrolujeme výskyt a stav kůrovců. Podle způsobu příprav rozeznáváme lapáky ležaté a stojaté. Kácíme-li ležaté lapáky opožděně, ponecháváme jim koruny, aby rychleji zavadly. Kácíme-li je včas nebo předčasně je lepší větve osekat. Stojaté lapáky připravujeme okroužkováním, tj. sloupnutím pásu kůry dokola kmene nebo vytvořením záseku až k jádrovému dřevu, popřípadě zkomolením koruny a odvětvením. Výhodnější jsou lapáky ležaté, které umožňují sledovat průběh nalétávání a vývoje kůrovců a ostatního podkorního a dřevokazného hmyzu. Průměry lapáků a jejich umístění se řídí druhem kůrovců, které chceme kontrolovat. Každý lapák musí být evidován a pod stálou kontrolou (Pfeffer 1961).
3.1.8. Požerky
Voroncov (1986) uvádí, že zvláštnosti stavby těla kůrovců těsně souvisejí s jejich způsobem života. Téměř po celý život žijí skrytě pod kůrou kmenů a větví stromů. Tam vytvářejí požerky, které mají tvar určitého obrazce. Pro každý druh kůrovce je zpravidla charakteristický požerek určitého tvaru. Požerky bývají jednoduché a složené. Jednoduché požerky sestávají z jednoho kanálku (matečné chodby), který vyhlodá samička, a bývají podélné nebo příčné. Složené požerky mají několik matečných chodeb a dělí se na hvězdicovité, se sklonem k podélnému nebo příčnému směru, a paprskovité. Každý druh kůrovce se vždy usídluje na určitém druhu dřeviny nebo na několika příbuzných druzích a obsazuje určitou část stromu.
Požerek je vyhlodaný systém chodeb určitého tvaru v kůře a běli, buď larvami, nebo dospělým hmyzem. Škůdci mohou vyhryzávat jednotlivé chodby nebo složité soustavy chodeb (Gusev et Rimskij-Korsakov 1953).
U dvou zmíněných rodů lze rozlišit jednak pod kůrou v lýku a částečně v běli zaříznutý požerek rodu Scolytus, který je rovnoběžný na osu kmene a larvové chodby jsou na něj kolmé a dále v běli probíhající požerky rodu Xyleborus. Na přítomnost kůrovců v dřevině upozorňují vypadávající drtinky, které mizí pouze po prudších deštích.
3.2. Rod Scolytus
Velikost 1,5–6,5 mm. Hlava je svrchu částečně viditelná. Čelo u samečků je ploché, často dlouze a hustě ochlupené. Čelo u samiček je vypouklé, lysé, nebo spoře ochlupené. Pouze u rodu Scolytus scolytus F. je hustě a krátce ochlupené. Tykadla se sedmičlenným bičíkem a dlouze oválnou paličkou, jejíž švy směřují ke středu paličky. Štít je po stranách zřetelně obroubený a svrchu tečkovaný. Tečky po stranách často splývají v podélné husté vrásky. Štít bývá obvykle temněji zbarven než krovky. Krovky velmi ploché, takže zadohruď a zadeček jsou dobře viditelné. Krovky jsou s řidšími či hustšími řádky teček. Zadeček má charakteristickou stavbu. První břišní článek navazuje normálně na zadohruď. Ostatní, počínaje druhým článkem, stoupají při pohledu ze strany šikmo vzhůru. Druhý, třetí a čtvrtý článek má u některých zástupců tohoto rodu uprostřed výrůstky nebo hrbolky, které jsou důležitým druhovým znakem. Povrch zadečku bývá lysý, nebo i ochlupený. Vnější okraj předních holení je hladký, neozubený a na konci hákovitě dovnitř zahnutý. Většina druhů tohoto rodu žije monogamicky a hlodá buď svislé, nebo příčné matečné chodby, které zasahují do povrchu běli stromů. Samička po ukončení snůšky zatarasí svým tělem vchod do chodby. Všechny naše druhy se vyvíjejí pod kůrou listnáčů. Z Alžírska a ze Sibiře jsou známy druhy, které se vyvíjejí v jehličnanech. Brouci po vylíhnutí se záhy prohlodávají kůrou na povrch a zalézají do korun zdravých stromů, kde se zavrtávají do nejmladších letorostů, v nichž hlodají krátké chodbičky, při tom pohlavně dospívají. Teprve potom zakládají nové pokolení. Některé druhy přenášejí při zralostním žíru spory hub, které potom ohrožují takto infikované stromy (grafióza jilmu). Z Evropy a přilehlých oblastí je známo 29 druhů (Pfeffer 1989). Hůrka (2005) uvádí, že rod je u nás zastoupen 12 druhy, které se vyvíjejí v listnatých stromech.
Kolařík (2004) uvádí, že floeofágní podkorní hmyz, kam rod patří, hlodá v lýku bohatém na živiny a napadá oslabené, odumírající nebo čerstvě odumřelé dřeviny, patří sem většina fytopatologicky významných druhů kůrovců (např. Ips typhographus L.). I floeofágní hmyz přenáší velké množství různých hub, a to buď aktivně v mycetangiích nebo pasivně ve vnitřnostech a v různých záhybech kutikuly. Některé tyto houby mohou být symbiotické. Nejznámější druhy těchto hub patří mezi vřeckovýtrusé do řádu Ophiostomatales (rod Ophiostoma) a Microascales (rod Ceratocystis). Obecně bývají nazývány jako ophiostomatální houby a hrají různé ekologické role. Mohou tvořit přídatný zdroj potravy larev a dospělců nebo oslabovat napadenou dřevinu. Tyto houby zpravidla zvyšují koncentraci dusíku – byla naměřena až dvakrát větší koncentrace než ve zdravém lýku. Houby hrají důležitou roli v narušení obranného systému dřevin. Jedním typem obranného mechanismu dřevin je výron pryskyřice. Houba rostoucí kolem požerku zaroste pryskyřičné kanálky a kůrovec, který je svým žírem naruší, pak není uvězněn v pryskyřici. Houby mohou pomáhat rozkládat látky chránící dřevo (zejména terpeny) před žírem a měnit hmyzí hormony (např. agregační – přileť, na antiagregační – strom je již obsazen). Jsou známi kůrovci, kteří nežijí s druhy ophiostomatálních hub a nedostatek dusíku nahrazují hlodáním velmi dlouhých chodeb. Mezi ně patří většina z rodu Scolytus. Tito kůrovci nemají mycetangia a donedávna se myslelo, že houby které přenášejí pasivně, patří pouze mezi druhy běžné v okolním prostředí (např. rod Penicillium). Jsou však pravidelně spojeni s vřeckovýtrusými hyfomycety rodu Geosmithia. Ty tvoří bílý porost v požercích kůrovců (např. Scolytus rugulosus). Zásadní význam těchto hub pro kůrovce není znám a jejich úloha je zřejmě je druhotná. Kratší život floeofágních kůrovců oproti ambróziovým je přikládán jejich výživě lýkem, které je díky terpenům mírně toxické.
3.2.1. Scolytus mali
3.2.1.1. Živná dřevina
Hůrka (2005) uvádí, že žije pod kůrou větví a slabších kmenů různých druhů růžovitých (Rosaceae), zvláště ovocných stromů. Schwenke (1974) uvádí výskyt na ovocných stromech – hrušni, rodu Prunus, kdouloni, dále na jeřábu, hlohu, skalníku. Výjimečně na jilmu a lísce. Vysoký (1995) uvádí konkrétně hrušeň, jabloň, třešeň, slivoň, myrobalán, meruňka, broskvoň, jeřáb, hloh, hlohyně, skalník, kdouloň, jilm, jasan a akát. Pfeffer (1989) uvádí pouze hrušeň, rod Prunus s. l., jabloň, jeřáb, hloh a skalník. A konečně Hluchý et al. (1997) uvádí hostitelské rostliny – jabloně, slivoně, třešně a další, jak ovocné stromy, tak jilmy, akáty aj.
3.2.1.2. Imago
Je 3,0–4,5 mm velký (Vysoký 1995, Hluchý et al. 1997, Hůrka 2005), náš největší kůrovec na ovocných dřevinách (Pfeffer 1989). Od příbuzného Scolytus rugulosus se pozná dle nápadného lesku (Lanák et al. 1969, Pfeffer 1989, Vysoký 1995, Hůrka 2005). Je černý (Pfeffer 1989), krovky červenavě hnědé až černé, lesklé (Vysoký 1995, Hůrka 2005). Schwenke (1974) uvádí krovky často červené. Lanák et al. (1969) uvádí, že brouk je červenohnědý. Hluchý et al. (1997) uvádí brouk s černou hlavou a štítem a hnědými krovkami. U nevyzrálých jedinců se objevují i červenavé krovky (Pfeffer 1989).
Tykadla a nohy červenavě hnědé. Čelo samců ploché až ploše vypouklé, ozdobené hustě podélně splývajícími rýžkami. Okraj nad kusadly je řídce ochlupený a až mezi oči lehce promáčklý (Vysoký 1995). U samic je čelo zřetelněji vypouklé (Pfeffer 1989, Vysoký 1995). Štít o málo delší než širší, velmi jemně tečkovaný (Vysoký 1995). Krovky v rýhách jemně tečkované (Pfeffer 1989, Vysoký 1995), mezirýží lesklé s řadou jemných teček, které jsou poloviční než tečky na rýhách (Vysoký 1995), mezirýží jsou plochá (Pfeffer 1989). Poslední břišní článek u samečka je uprostřed mělce vyhloubený (Pfeffer 1989). Články zadečku jsou rovnoměrně zkoseny, u samců je na posledním článku jamka (Vysoký 1995). Sameček se liší od samičky krátkým a řídkým ochlupením čela. Samička má čelo vypouklé a téměř lysé (Pfeffer 1989).
3.2.1.3. Bionomie
Vyvíjí se pod kůrou ovocných dřevin, nejčastěji kmenů a silných větví (Lanák et al. 1969, Vysoký 1995), často se Scolytus rugulosus s nímž má shodný vývoj, ale Scolytus rugulosus je ve slabších větvích a kmíncích. Požerky obou druhů se mohou proplétat. Páry jsou monogamní (Vysoký 1995).
Ve střední Evropě má dvě generace do roka (Lanák et al. 1969, Schwenke 1974, Pfeffer 1989, Vysoký 1995, Hluchý et al. 1997, Hůrka 2005). Vzácností není sesterské pokolení (Vysoký 1995). Rojí se v V–VI a VIII–IX (Schwenke 1974, Vysoký 1995, Hluchý et al. 1997), druhá generace přezimuje v larválním stádiu (Schwenke 1974).
Celý vývoj trvá 11–12 týdnů (Lanák et al. 1969, Vysoký 1995, Hluchý et al. 1997). Přezimovat mohou larvy a kukly (Lanák et al. 1969, Vysoký 1995, Hluchý et al. 1997). Má-li škůdce jen jedno pokolení, pak přezimuje brouk v chodbičce (Lanák et al. 1969, Vysoký 1995).
Žír probíhá v kůře živých stromů. Často je uváděn jako škůdce ovocných dřevin spolu s bělokazem Scolytus rugulosus, v listnatých lesích patří k řídce se vyskytujícím druhům. Napadá hlavně stromy nemocné, chřadnoucí po suchých letech, poškozené mrazy, okusem nebo onemocnělé rakovinou (Lanák et al. 1969, Kutina 1983, Vysoký 1995, Hluchý et al. 1997). Vyskytuje se i v pařezech a pokácených nebo polámaných kmenech. Ohniskem se mohou stát opuštěné zahrady, odkud se brouci rozletí do okolí. V případě přemnožení napadá i zdravé a mladé štěpy (Vysoký 1995).
3.2.1.4. Vajíčko
Vajíčka oválná, bílá o velikosti 0,8 x 0,4 mm (Vysoký 1995), měkká, stočená (Lanák et al. 1969).
3.2.1.5. Larva
Larvy jsou eucephalní a apodní (beznohé). Maxily jsou catopoidní. Tykadla jsou krátká, pouze s jediným kratičkým článkem (Kalina 1999). Larva bílá, měkká, velikosti asi 4 mm, ventrálně stočená s četnými záhyby, s hojnými mikroskopickými chloupky (Pfeffer 1989, Vysoký 1995). Někdy slabě narůžovělá, neboť prosvítá zažívací ústrojí (Pfeffer 1989). Hlava žlutohnědá až hnědá (Vysoký 1995), chitinizovaná.
3.2.1.6. Kukla
Kukla volná (pupa libera). Kuklí se v běli (Kalina 1999, Vysoký 1995, Lanák et al. 1969). V posledním stádiu před líhnutím silně chitinizovaná, všechny orgány patrné, křídla zřetelné (Vysoký 1995, Lanák et al. 1969).
3.2.1.7. Příznaky napadení
Na větvích a kmeni způsobuje zakrňování a odumírání stromů s vyžranými chodbami pod kůrou (Nienhaus et al. 1998). Na kmenu jsou zvenčí viditelné závrty, z nichž vypadává vyhlodaná drť, ukazují na napadení tímto kůrovcem. Na napadených třešních, višních a švestkách se objevuje klejotok z vyhlodaných otvorů (Lanák et al. 1969, Vysoký 1995, Hluchý et al. 1997). Činností tohoto škůdce stromy prosychají a větve během vegetace vadnou (Lanák et al. 1969). Strom prosychá nejprve v koruně a příznakem je vadnutí, žloutnutí a opadávání listí i plodů ve vegetační době (Vysoký 1995).
3.2.1.8. Popis požerku
Pod kůrou kmenů a tlustých větví chřadnoucích ovocných stromů (Kutina 1983, Hluchý et al. 1997) jsou pravidelné požerky (Lanák et al. 1969). Matečná chodba je svislá (Gusev et Rimskij-Korsakov 1953, Lanák et al. 1969, Pfeffer 1989), jednoramenná (Lanák et al. 1969, Schwenke 1974). Začíná snubní komůrkou v lýkové části kmene (Lanák et al. 1969) s rozšířením na začátku. Široká je 1,6 mm (Vysoký 1995) až 2 mm i více, (Gusev et Rimskij-Korsakov 1953, Lanák et al. 1969, Hluchý et al. 1997). Matečná chodba 2 (Schwenke 1974, Vysoký 1995) až 7 cm (Gusev et Rimskij-Korsakov 1953) nebo až 12 cm (Lanák et al. 1969, Kutina 1983, Hluchý et al. 1997) dlouhá. Po každé straně mateřské chodby je 30–60 larvových chodeb (Lanák et al. 1969, Kutina 1983, Vysoký 1995), které jsou zpočátku kolmé nebo mírně skloněné k mateřské chodbě (Gusev et Rimskij-Korsakov 1953, Lanák et al. 1969), jsou vlnité (Vysoký 1995), spodní se ohýbají podél kmene v tlusté kůře (Gusev et Rimskij-Korsakov 1953). Mateřské i larvové chodby zasahují do běle a hloubka otisku chodeb v běli je závislá na vlhkosti kmene, na suchých kmenech je otisk ostřejší – chodby zasahují do dřeva podle svěžesti kmene (Lanák et al. 1969, Vysoký 1995). Na koncích larvových chodeb jsou lůžka kukel (Lanák et al. 1969) s kukelní komůrkou v běli (Vysoký 1995). Napadení nikdy nejde hluboko do dřeva (Hluchý et al. 1997).
3.2.1.9. Parazité a predátoři
Bělokazi mají řadu druhů blanokřídlých parazitoidů (Hluchý et al. 1997).
Vysoký (1995) uvádí tyto druhy parazitů – Eurytoma roseae Nees, Microplectron fuscipennis Zett., Doryctes pomarius Reinh., Elachistus leucogramma R., Microplectron leucograma R., Pteromalus sp., Rhaphitelus maculatus Walk., Plectonchus corontus Fuchs, Scinura mali Fuchs. Predátory kůrovců jsou draví zástupci nadčeledi vosy (Vespoidea).
3.2.1.10. Rozšíření
Rozšířen v celé Evropě po Kavkaz a Ural (Schwenke 1974), jižní, střední a východní Evropa. V ČR je rozšířen od nejnižších poloh až po úpatí hor, nejvíce v pahorkatinách (Vysoký 1995, Pfeffer 1989).
3.2.2. Scolytus rugulosus
3.2.2.1. Živná dřevina
Schwenke (1974) uvádí výskyt na ovocných stromech a planých Rosaceae, dále znám na Betula, Alnus, Ulmus a Corylus avellana L. Pfeffer (1989) uvádí, že se vyvíjí v různých druzích rodu Pyrus, Prunus, Malus, Sorbus, Crataegus, Rosa aj. Gusev et Rimskij-Korsakov (1953) uvádí Pyrus communis L., Malus sylvestris Mill., Sorbus aucuparia L., Prunus armeniaca L., Prunus domestica L., Prunus avium L., Prunus cerasus L. A konečně Hluchý et al. (1997) uvádí jako hostitelské rostliny jabloně, slivoně, třešně a další, jak ovocné stromy, tak jilmy, akáty aj.
3.2.2.2. Imago
V elikost 1,4–3,2 mm (Pfeffer 1989), 1,8–2,5 mm (Lanák et al. 1969), 1,8–3,0 mm (Schwenke 1974) dlouhý a černý (Lanák et al. 1969, Schwenke 1974) s tmavě hnědými až černými krovkami (Schwenke 1974). Pfeffer (1989) uvádí zbarvení tmavohnědé až černé. Okraje štítu a konec krovek jsou hnědavě červené. Štít hustě a hrubě tečkovaný. Po stranách štítu jsou tečky protáhlé a splývající. Krovky hustě, hrubě a v hustých rýhách tečkované. Mezirýží jsou úzká a taktéž silně tečkovaná. Čelo samečka je spoře a krátce ochlupené, čelo samičky vypouklé a velmi krátce a velmi spoře ochlupené. Šev mezi prvním a druhým břišním článkem je nezřetelný. Jde o neobyčejně proměnlivý druh co do velikosti, barvy i struktury těla.
Ve velikosti a barvě se sice autoři úplně neshodují, avšak shodují se v tom, že jde vždy o brouka menšího než příbuzný Scolytus mali.
3.2.2.3. Bionomie
Často se Scolytus mali s nímž má shodný vývoj (Kutina 1983, Vysoký 1995, Hluchý et al. 1997), ale Scolytus rugulosus je ve slabších větvích a kmíncích. Požerky se mohou proplétat. Páry jsou monogamní (Vysoký 1995).
Ve střední Evropě obvykle dvě generace do roka (Lanák et al. 1969, Schwenke 1974, Pfeffer 1989, Hluchý et al. 1997). Rojí se v V–VI a VIII–IX (Hluchý et al. 1997), druhá generace přezimuje v larválním stádiu (Schwenke 1974).
Celý vývoj trvá 9–10 (Lanák et al. 1969), 11–12 týdnů (Hluchý et al. 1997). Přezimovat mohou larvy a kukly (Lanák et al. 1969, Hluchý et al. 1997). Má-li škůdce jen jedno pokolení, pak přezimuje brouk v chodbičce (Lanák et al. 1969).
Žír probíhá v kůře živých stromů. Často uváděn jako škůdce ovocných dřevin spolu s bělokazem Scolytus mali. Napadá hlavně stromy nemocné, chřadnoucí po suchých letech, poškozené mrazy, okusem nebo onemocnělé rakovinou (Lanák et al. 1969, Kutina 1983, Hluchý et al. 1997). Výskyt tohoto bělokaza je větší v blízkosti lesa. Brouci, než dospějí, ožírají pupeny nebo kůru v okolí pupenů (Lanák et al. 1969).
3.2.2.4. Vajíčko
Podobné Scolytus mali.
3.2.2.5. Larva
Podobná Scolytus mali.
3.2.2.6. Kukla
Podobná Scolytus mali.
3.2.2.7. Příznaky napadení
Shodné se Scolytus mali, vyskytuje se však hlavně na mladých stromcích po výsadbě (Kutina 1983) a tenčím materiálu.
3.2.2.8. Popis požerku
Na větvích a kmenech s tenkou kůrou ovocných stromů jsou požerky (Gusev et Rimskij-Korsakov 1953, Hluchý et al. 1997). Matečná chodba je svislá (Gusev et Rimskij-Korsakov 1953, Lanák et al. 1969, Pfeffer 1989), jednoramenná (Lanák et al. 1969, Schwenke 1974). Začíná snubní komůrkou v lýkové části kmene (Lanák et al. 1969). Užší než 2 mm, konkrétně 1,2–1,8 mm široké, bez rozšíření na začátku (Gusev et Rimskij-Korsakov 1953, Lanák et al. 1969, Hluchý et al. 1997). Matečná chodba krátká 1 až 3 cm dlouhá (Gusev et Rimskij-Korsakov 1953, Schwenke 1974) až 4 cm dlouhá (Lanák et al. 1969). Po stranách husté larvové chodby (Pfeffer 1989), klikaté v různých směrech pěkně otisknuté do dřeva, hadovité (Gusev et Rimskij-Korsakov 1953). Larvové chodby nemají tak charakteristické rozvětvení jako u Scolytus mali a na mateřské chodbě je jich po každé straně podstatně méně, konkrétně 10–20 (Lanák et al. 1969, Kutina 1983). Chodby mnohem kratší než u Scolytus mali (Kutina 1983). Na koncích larvových chodeb jsou lůžka kukel s kukelní komůrkou v běli (Gusev et Rimskij-Korsakov 1953). Napadení nikdy nejde hluboko do dřeva (Hluchý et al. 1997).
3.2.2.9. Parazité
Bělokazi mají řadu druhů blanokřídlých parazitoidů (Hluchý et al. 1997). Spektrum bude zřejmě obdobné jako u předchozího druhu.
3.2.2.10. Rozšíření
V střední a jižní Evropě, jihu severní Evropy, Rusko po Kavkaz, Krym (Schwenke 1974).
3.3. Rod Xyleborus
Zástupci tohoto rodu jsou význační svým pohlavním dimorfismem (Voroncov 1986, Pfeffer 1989, Hůrka 2005). Samečci jsou vždy menší (Pfeffer 1989, Hůrka 2005) o velikosti 0,3–3,0 mm, se zakrnělými křídly, žlutohnědí až tmavohnědí. Okřídlené samičky jsou 2,0–4,0 mm velké, mahagonově hnědé až černohnědé a odstále ochlupené (Pfeffer 1989, Hůrka 2005). Tykadla s krátkým pětičetným bičíkem a krátce oválnou, na vnitřní straně tupě zakončenou, tříčlennou paličkou. Štít u samiček je polokulovitý, nebo krátce válcovitý a vpředu zřetelně hrbolkovaný, vzadu pak silněji či jemněji tečkovaný, nebo hladký či šagrenovaný. Samečci mají štít vpředu neznatelně hrbolkovaný, plochý nebo kápovitě dopředu protažený. Krovky u samiček jsou víceméně válcovité, vzadu rovnoměrně vyklenuté, obvykle s malými hrbolky. Samečci mají krovky válcovité až kulovité, nebo zploštělé, v řádcích řidčeji a jemněji tečkované než u samiček. Zadní část krovek podobně hrbolkovaná jako krovky samiček. Drtinky u závrtových otvorů jsou práškovité. Samičky hlodají v běli stromů, řidčeji v lýku silné borky, různě utvářené matečné chodby. Vylíhlé larvy se živí podhoubím ambróziových hub (Pfeffer 1989, Hůrka 2005), konkrétně Ambrosiella hartigii Batra (syn. Monilia candida Hart.) (Pfeffer 1989, Six 2003 et Harrington 2005 in Šrůtka 2006), které pokrývají stěny matečných chodeb. Pozdě v létě se líhnou noví brouci, převládají samičky. Samečci se objevují v chodbách po vylíhnutí jednotlivě. U druhu Xyleborus cryptographus Ratzb. v tisícinách, u ostatních druhů ve více procentech z vylíhlých jedinců. Některé druhy mohou být technickými škůdci dřeva. V Evropě se vyskytuje 7 druhů domácích a jeden druh zavlečený z východní Asie (Pfeffer 1989).
Na světě žije v různých klimatických pásech více než 1300 druhů kůrovcovitých (Scolytinae) z rodu Xyleborus Eichh. (Urban 2004). Rod má v ČR a SR 6 zástupců. Vyvíjejí se ve dřevě nejrůznějších listnáčů (Hůrka 2005), na kterých jsou velmi rozšíření (Voroncov 1986), 1 druh v borovicích (Hůrka 2005). Urban (2004) uvádí, že ve fauně ČR jsou tito dřevokazní kůrovci zastoupeni 8 druhy, z nichž 7 se vyvíjí v běli listnáčů a 1 v běli jehličnanů. K nejvýznamnějším z nich patří drtník prostřední (X. monographus F.), drtník všežravý (Xyleborus saxeseni) a drtník ovocný (Xyleborus dispar). Požerky jednotlivých druhů drtníků se vzájemně značně liší (Voroncov 1986).
Chodby ambróziových brouků mají délku jen několik málo centimetrů, jsou duté, nevyplněné drtinkami a všude stejně široké, protože larva nebo dospělec při sklízení své zahrádky leze v obou směrech. Nově vylíhlí brouci opouštějí dřevo otvorem, který vyhlodají dospělci. Ambróziový brouci nehynou po ukončení rozmnožování, jako je tomu u floeofágních kůrovců. Dále spásají houbová mycelia a zatím neznámým způsobem udržují monokulturu ambróziové houby. Když dospělci ambróziových kůrovců zahynou, larvy dál také žít nemohou, protože neumějí udržovat čistotu houbové zahrádky. Také druhy rodu Fusarium jsou považovány za ambróziové houby kůrovců z tribu Xyleborini (Kolařík 2004).
3.3.1. Xyleborus dispar
3.3.1.1. Živná dřevina
Je polyfágem na listnáčích (Schwenke 1974, Voroncov 1986, Kalina 2004). Vyvíjí se v běli všech listnáčů (Pfeffer 1989). Vývoj téměř ve všech listnatých dřevinách, nejvíce v dubu, buku, olši, kaštanu, ořešáku, habru, bříze, topolech, vrbě, jasanu, klenu, akátu, vinné révě, jeřábu, lísce, lípě, javoru, bezu a v ovocných stromech (např. jabloň, hrušeň, slivoň, střemcha) (Novák et al. 1974, Vysoký 1995, Hluchý et al. 1997), pozorován i v jehličnanech (Pfeffer 1954 in Vysoký 1995), v zeravu a borovici (Miller 1956 in Vysoký 1995, Schwenke 1974, Urban 2004), jalovci (Schwenke 1974, Urban 2004), smrku, jedli, douglasce (Urban 2004). Druh se vyvíjí ve větvích i kmenech téměř všech listnatých stromů. Je významným technickým škůdcem dřeva (Hůrka 2005). Gusev et Rimskij-Korsakov (1953) uvádí jabloň lesní a jiné druhy, líska obecná, meruňka obecná, ořešák vlašský, třešeň ptačí a višeň obecná.
Nejhojnější je na dubu, buku, olši, javoru, jabloni, hrušni, švestce a meruňce. Vyskytuje se však např. i na bříze, jasanu, kaštanovníku, ořešáku, habru, lípě, topolu, vrbě, platanu, akátu, jeřábu, lísce, střemše, bezu, břečťanu, zimostrázu, kdouli, révě vinné, višni, třešni aj. při přemnožení na Moravě byl jeho výskyt zaznamenán hlavně na dubu, buku, jabloni, meruňce, hrušni, slívě a švestce. V lesní školce na Hádech působil hynutí 1 až 2 m vysokých odrostků jírovce maďalu (Aesculus hippocastanum L.) a svitelu latnatého (Koelreuteria paniculata Laxm.) (Urban 2004).
3.3.1.2. Imago
U tohoto druhu je pohlavní dimorfismus. Sameček 1,8–2,1 mm (Lanák et al. 1969, Novák et al. 1974, Pfeffer 1989, Vysoký 1995, Hůrka 2005), 1,8–2,4 mm (Schwenke 1974, Urban 2004) 1,5–2,5 mm (Hluchý et al. 1997), světlehnědě (Schwenke 1974, Urban 2004) až tmavohnědě zbarvený (Pfeffer 1989, Urban 2004), matný (Urban 2004), bezkřídlý – křídla zakrnělá (Pfeffer 1989, Vysoký 1995, Urban 2004), tělo kulovité, krovky tak dlouhé jak široké (Pfeffer 1989, Urban 2004), jsou silně vypuklé, takže z boku je tento kůrovec charakteristicky sražený (Novák et al. 1974). Jsou v řádkách hrubě tečkované, štít plochý, svrchu jemně tečkovaný, odstále dlouze chloupkovaný. Přední okraj slabě hrbolkovaný (Novák et al. 1974, Vysoký 1995). Krátké a zepředu zploštělé tělo je morfologicky adaptováno ke kopulaci v matečných chodbách (Urban 2004). Samička 3,0–3,6 mm (Novák et al. 1974, Vysoký 1995), 3,0–3,8 mm (Schwenke 1974, Urban 2004), 3,2–3,6 mm (Pfeffer 1989, Hůrka 2005), 3,0–3,5 mm (Lanák et al. 1969, Hluchý et al. 1997), tělo hnědočerné až černé (Schwenke 1974, Pfeffer 1989, Urban 2004), válcovité (Novák et al. 1974, Pfeffer 1989, Vysoký 1995), křídla dobře vyvinuta (Pfeffer 1989, Vysoký 1995, Urban 2004), krovky široce válcovité, hrubě a zřetelně v rýhách tečkované (Novák et al. 1974, Pfeffer 1989, Vysoký 1995, Urban 2004), lysé (Pfeffer 1989, Vysoký 1995, Urban 2004), kryjí zadohruď i zadeček (Vysoký 1995). Štít vpředu hrbolkovaný (Novák et al. 1974, Vysoký 1995, Pfeffer 1989), po stranách neobroubený (Vysoký 1995), vzadu šagrenovaný, matný a velmi jemně a řídce tečkovaný (Pfeffer 1989).
U obou pohlaví nožky a tykadla žlutavá (Novák et al. 1974, Vysoký 1995, Urban 2004). Štít dlouhý jako široký (Vysoký 1995) a tělo odstále chloupkované (Urban 2004).
3.3.1.3. Bionomie
Rauleder (2004) uvádí, že rojení začíná na jaře, když teploty dosáhnou 18°C a více. To odpovídá konci III, až prvnímu týdnu v IV. Rojení pokračuje až do začátku VII. Ostatní autoři uvádí rojení samiček od 2. poloviny IV až V (Urban 2004), ev. IV až V (Lanák et al. 1969, Schwenke 1974, Vysoký 1995, Kalina 2004), ev. V až VI (Novák et al. 1974, Hluchý et al. 1997) náletem na čerstvé pařezy, odumírající stromy, na pokácené neodkorněné dříví (Novák et al. 1974, Vysoký 1995) a na větve (Hluchý et al. 1997). Má jedno pokolení a přezimuje imago (Lanák et al. 1969, Schwenke 1974, Vysoký 1995, Hluchý et al. 1997, Kalina 2004). Brouci přezimují z největší části v chodbách natlačení těsně vedle sebe, pouze menší část samiček v létě vyletuje a přezimuje jinde (Lanák et al. 1969, Kalina 2004), někdy přezimují zvlášť samečkové a zvlášť samičky (Lanák et al. 1969). Oplození probíhá v chodbách, samečkové jsou neschopní letu a jsou v daleko menším počtu než samičky (Kalina 2004). Poměr pohlaví (♂:♀) kolísá 1:4 – 1:44, obvykle 1:5 (Vysoký 1995), 1:6 až 7 (Novák et al. 1974), obvykle 1:5 až 7 (Urban 2004).
Samička klade vajíčka postupně, jak vrtá chodbičky, do hromádek asi po 6 kusech (Vysoký 1995), skupiny vajíček přikrývá vlhkou dřevní drtí, čímž se vytvářejí vhodné podmínky pro růst výživných hub (Lanák et al. 1969). Celkem během 1,5 až 2 měsíců vykladou asi 30–40 vajíček na samičku (Vysoký 1995, Urban 2004). Poslední vykladená vajíčka je možno v chodbách zastihnout ještě koncem VI (Urban 2004).
Po několika dnech (Miller 1956 in Vysoký 1995), ev. po 14 dnech (Vysoký 1995) se z vajíček líhnou larvy. Vylíhlé larvičky, jakož i rodičovští brouci – a nakonec i mladí, noví dospělci se neživí dřevem, nýbrž podhoubím houby Endomyces pěstovaným na stěnách chodeb (Novák et al. 1974), tzv. ambrozií (Vysoký 1995). Dle Pfeffera (1955 in Vysoký 1995) jde konkrétně o Leptographium. Spory hub bývají zaneseny do chodeb dospělci. Dříve se autoři domnívali, že je chovají ve středním střevě (Miller 1956 in Vysoký 1995), dle Blattného (1956 in Vysoký 1995) udržují samice spory v žaludku a výtrusy přenášejí kusadly. Nejnovější poznatky však jasně hovoří o přenosu hub v mycangiích (viz 3.1.4.) (Batra 1963 in Šrůtka 2006) a podhoubí tvoří houba Ambriosiella hartigii Batra (Urban 2004, Šrůtka 2006).
Na množství houby závisí úspěšný vývoj tohoto druhu. Samice je schopna udržovat ucpáváním a větráním chodeb vhodné klima pro pěstování houby na stěnách chodeb (Vysoký 1995, Urban 2004). Stěny chodeb brzy porůstá smetanově bílé mycelium, které samičky okusují, a tím ho udržují v optimálním stavu pro svou výživu (Urban 2004).
Larvy se vyvíjejí postupně (Hluchý et al. 1997). Žijí volně v matečných chodbách a žerou asi 5–6 týdnů. Vývoj dokončují v červenci a hned se kuklí (Novák et al. 1974, Vysoký 1995). V důsledku pomalého hloubení chodeb a dlouhého období kladení se v nejdříve zhotovených chodbách vyskytují poloodrostlé larvy, zatímco mateřští brouci teprve dokončují hloubení matečných chodeb a kladení vajíček. Larvy se v chodbách vyskytují skupinovitě, a když dorostou, bývají v chodbách nahloučeny těsně jedna za druhou a svými těly vyplňují skoro celý příčný profil chodeb. Bílými, lesklými těly ostře kontrastují se stěnami chodeb, které postupně v důsledku úhynu podhoubí červenají, hnědnou, až černají (Urban 2004).
Během vývoje larev se samička zdržuje u otvoru chodby a stará se o čistotu v chodbičkách – odstraňuje výkaly. Většina samiček následně hyne. Dorostlé larvy se kuklí v chodbičkách (Lanák et al. 1969, Vysoký 1995, Hluchý et al. 1997) asi po 7 až 10 dnech (Novák et al. 1974, Vysoký 1995), po 10 až 14 dnech se z nich líhnou brouci (Lanák et al. 1969). Mladí brouci žijí rovněž v chodbičkách, kde při zralostním žíru spásají podhoubí. Během několika dnů pohlavně dospívají a vyhledávají mladé sesterské samičky, s nimiž se v chodbách páří. I přes malý počet samečků jsou do podzimu oplodněny všechny samičky (Urban 2004).
Přezimují mladí brouci v chodbách (Schwenke 1974, Urban 2004). Část samiček dává vznik tzv. sesterskému potomstvu, které samičky zakládají v VII až VIII (Lanák et al. 1969, Vysoký 1995). Letní závrty na mladých listnáčích, popř. na větvích starších stromů se připisují rojení a opakované snůšce rodičovských brouků, kteří opouštějí kolonie od doby, kdy se v nich objeví první kukly a mladí brouci (Novák et al. 1974). Dle Pfeffera (1955 in Vysoký 1995) druhé pokolení není bezpečně prokázáno. Dnes se autoři přiklání k názoru, že drtník ovocný je typicky univoltinní druh (druh s jednoletým vývojem). Za příznivého počasí někdy mladé samičky požerky předčasně opouštějí a zalézají do štěrbin kůry, nebo se zavrtávají do mladých kmínků a větví, kde v hloubce kolem 5 mm zimují. Takovéto neobvyklé chování bylo považováno za úživný žír předcházející tvorbě druhé generace nebo za sesterské pokolení jarního náletu (Urban 2004).
Přes zimu jsou dospělci seřazeni v chodbách těsně za sebou a hlavou orientováni ke slepým koncům chodeb (dovnitř dřeva). Samečci většinou přežívají až do jara, kdy se opakovaně páří se samičkami, načež pak chodby opouštějí a hynou. Samička většinou umírá až po vylíhnutí mladých brouků (Urban 2004). Vylíhlé brouky nalezneme v chodbách v řadě za sebou otočené hlavou do nitra kmene. Samečci bývají obvykle pohromadě. V těchto chodbách probíhá také páření a oplodněné samičky se rozlétají zakládat nové pokolení. Celý vývoj trvá 10–11 týdnů (Vysoký 1995).
Na rozdíl od podkorních druhů kůrovců si drtníci nezhotovují výletové otvory. Požerky opouštějí vstupními chodbami a to pozpátku couváním. Role samečků oplozením samiček končí a veškerou péči o založení nové generace a její zdárný vývoj přebírají samičky (Urban 2004).
3.3.1.4. Vajíčko
Vajíčko bílé a měkké, 0,8 – 0,9 x 0,4 mm, oválné (Vysoký 1995).
3.3.1.5. Larva
Larvy jsou eucephalní a apodní – beznohé (Kalina 1999), avšak dost pohyblivé (Vysoký 1995). Maxily jsou catopoidní. Tykadla jsou krátká, pouze s jediným kratičkým článkem (Kalina 1999). Larva bílá, měkká, velikosti asi 4 mm, ventrálně stočená. Hlava hnědavá (Vysoký 1995).
3.3.1.6. Kukla
Kukla volná (pupa libera). Kuklí se ve dřevě. Samičí kukla má dva páry křídel, u samečků pouze jeden (Lanák et al. 1969, Vysoký 1995). Kukla je bílá, až před líhnutím se zbarvuje hnědě (Vysoký 1995).
3.3.1.7. Příznaky napadení
Přítomnost drtníka ovocného ve dřevě signalizují závrtové otvory o průměru kolem 1,7 mm a z nich vypadávající práškové drtinky (Novák et al. 1974, Pfeffer 1989, Urban 2004), žlutě zbarvené, podle nichž byl drtník v češtině pojmenován (Vysoký 1995). Kutina (1983) uvádí ne zcela přesně, že kruhovité závrtové otvory jsou na 2–3letém dřevě jednotlivé. Okolí závrtů na některých dřevinách (např. na dubu) tmavne vlivem výronu mízy (Vysoký 1995). To je spíše způsobeno působením symbiotických hub kůrovců. U nás patří k největším potenciálním škůdcům v ovocnictví (Vysoký 1995). Tenčí větve usychají nad místy, kde vrtali drtníci. Napadnou-li drtníci silnější větve, následují je kůrovci, jejichž vývoj začíná v místě napadení drtníky, a různé houby. Při silnějším napadení strom usychá, slabší napadení strom může přežít, ale je oslaben a živoří (Lanák et al. 1969). Napadení drtníky však může vést k usychání celých větví, případně i celých stromů (Hluchý et al. 1997). Determinace škůdce podle vnějších příznaků nebo hloubkové analýzy požerků je však vždy orientační, a proto je nutné ji pojistit determinací dospělců (Urban 2004). Významný především jako technický škůdce (Kalina 2004).
3.3.1.8. Popis požerku
Požerek drtníka ovocného je poměrně specifický (Voroncov 1987). Matečná chodba probíhá v běli kmenů (Gusev et Rimskij-Korsakov 1953, Pfeffer 1989) a larvální chodby uvnitř dřeva v podobě složitého obrazce (Gusev et Rimskij-Korsakov 1953). V prasklinách kůry, úžlabích větví a jiných nerovnostech se kolmo zahlubují do kůry a ve dřevě pak hloubí přibližně radiální vstupní chodbu. Ta je 0,3 (Urban 2004) až 6 cm dlouhá (Pfeffer 1989, Vysoký 1995, Novák et al. 1974) dle tloušťky dřeva a na příčném řezu kruhovitá – 1,5 až 1,9 mm (Pfeffer 1989, Vysoký 1995, Urban 2004) široká. Od vstupní chodby samičky dále hlodají matečné chodby prvního řádu – primární chodby, které probíhají na obě strany zhruba ve směru letokruhů, ve slabším materiálu často na jednu stranu. Od nich pak v nepravidelných odstupech vyžírají různě dlouhé a různě početné matečné chodby druhého řádu – sekundární chodby, které jsou alespoň v koncových úsecích rovnoběžné s dřevními vlákny (Miller 1956 in Vysoký 1995, Lanák et al. 1969, Pfeffer 1989, Kalina 2004, Urban 2004) dlouhé až 3 cm (Voroncov 1987). V tenkých kmíncích a větvích matečné chodby často vybíhají z krátké vstupní chodby, a to v různé celkové délky obvykle 2 až 4 cm. Všechny chodby v požerku mají stejný průměr kolem 1,7 mm a zhotovuje je jedna samička. Ta chodby průběžně čistí, a proto její přítomnost ve dřevě prozrazují jemné žlutobílé práškovité drtinky s trusem, které se často hromadí ve štěrbinách na povrchu kůry (Urban 2004). Délka chodeb je variabilní dle tloušťky napadeného dřeva (Vysoký 1995). Stejnými vnějšími symptomy napadení se vyznačuje dřevokaz bukový (Xyloterus domesticus L.). Jeho matečné chodby jsou však delší a probíhají většinou v příčné rovině. Ve směru dřevních vláken z nich odbočují nahoru a dolů kratičké larvové chodbičky o délce kolem 5 mm. Dřevokaz bukový poškozuje, velmi zřídka ovocné stromy (Urban 2004).
3.3.1.9. Parazité a predátoři
Ve srovnání s bělokazy je v případě drtníků známo podstatně méně jejich parazitoidů (Hluchý et al. 1997). Predátoři – Rhizophagus aeneus Richt. (ale pouze na olši), Agnathus decoratus Germ., Salpingus planirostris F., Colydium filiforme F., Corticeus bicolor Ol.
Parazité – Pseudopimla anisandri Fahr., Perniphora robusta Rusch., Rhabditis juglandicola Fuchs, Anmguillulina pinguicauda Fuchs (Vysoký 1995).
3.3.1.10. Rozšíření
Evropa, severní Afrika, Krym, Kavkaz, Přední Asie, až po západní Sibiř, Kanada, USA. V ČR všude mimo horské lesy. Nejhojněji v luzích a lesostepi od 300 do 700 m n. m., v parcích i zahradách (Vysoký 1995). Schwenke (1974) uvádí, že je ve střední a jižní Evropě, jihu severní Evropy, východní Evropa po Sibiř a severní Afrika. Pfeffer (1989) uvádí, že se hojně vyskytuje v nížinách a v pahorkatinách. A konečně Urban (2004) uvádí, že drtník ovocný se vyskytuje v převážné části Evropy (od Portugalska a Itálie až do střední Skandinávie), v severní Africe, západní Sibiři a malé Asii. V ČR je hojný v řídkých listnatých případně smíšených lesích různého věku, na okrajích pasek, v parkových a alejových výsadbách a ovocných sadech, a to hlavně v nížinách a pahorkatinách. V nižších horských polohách je řidší, a schází v horských jehličnatých lesech.
3.3.2 Xyleborus saxeseni
3.3.2.1. Živná dřevina
Extremně polyfágní druh s výraznou preferencí pro listnáče, vývoj však zaznamenán i na jehličnanech (Kalina 2004, Urban 2004). Duby, buky, lípy, korkovník, vrba, jilmy, javor, habry, břízy, olše, topoly, kaštan, líska, ořešák, jasany, jeřáb, ovocné stromy (např. slivoň, jabloň, hrušeň), borovice, smrk, jedle, modřín (Vysoký 1995). Jabloň lesní a jiné druhy, meruňka obecná, ořešák vlašský, slivoň domácí, třešeň ptačí (Gusev et Rimskij-Korsakov 1953). Napadá mnohé listnaté dřeviny (dub, olši, lísku aj.) a na Dálném Východě i jehličnany (Voroncov 1986), znám je z jehličnatých – borovic, smrků, modřínů, cedrů, jedlí nebo jedlovců (Schwenke 1974).
3.3.2.2. Imago
Jde o druh s pohlavním dimorfismem. Je to nejmenší 1,5 až 2,4 mm velký domácí drtník (Urban 2004). Sameček (1,5) 1,8–2 mm, světlehnědý (Schwenke 1974, Vysoký 1995). Zadní části štítu a přední část krovek jsou kratičce ochlupeny. Šev, třetí a páté mezirýží na zadní části krovek s tupými hrbolky (Pfeffer 1989). Má zakrnělá křídla. Samička 2,0–2,4 mm (Schwenke 1974, Kutina 1983, Pfeffer 1989, Vysoký 1995), tmavohnědá (Schwenke 1974, Vysoký 1995), hlava a štít světlejší (Vysoký 1995). Je úzká a válcovitá, štít vpředu jemně hrbolkovaný. Štít velmi jemně a řídce tečkovaný. Krovky v rýhách jemně tečkované, válcovité, lesklé a krátce ochlupené (Vysoký 1995). Druhé nehrbolkované mezirýží je slabě prohloubené. Hrbolky na švu a na třetím mezirýží jsou tupé (Pfeffer 1989). Štítek nezřetelný (Pfeffer 1989, Vysoký 1995).
Nohy a tykadla jsou u obou pohlaví světle žlutá (Vysoký 1995).
3.3.2.3. Bionomie
Obvykle dvě pokolení do roka (Vysoký 1995, Kalina 2004). Rojení v IV a V (Schwenke 1974, Vysoký 1995, Kalina 2004). V létě v VII a VIII (Vysoký 1995, Kalina 2004), z tohoto pokolení přezimují různá vývojová stádia (Vysoký 1995). Zpravidla čerství brouci v chodbách, zřídka larvy nebo kukly (Kalina 2004). Hlodá v běli matečnou chodbu (Pfeffer 1989). Samičky nehlodají vždy matečné chodby, ale žijí v chodbách předchozího pokolení, kde jen zvětšují požerek, vzniká kolonie různého stáří. Napadá dřevo poražených a neodkorněných stromů, nemocné stromy, čerstvé pařezy, kde doprovází žír jiných druhů hmyzu, dále i stromy zasažené bleskem. Samička klade vajíčka s přestávkami, během kterých příjmá potravu (Voroncov 1986). Vývoj nepravidelný, lze zastihnout ve stejné době všechna vývojová stádia (Voroncov 1986, Vysoký 1995). Mladí brouci zůstávají dlouho v chodbách, souvisí to s tím, že v chodbách samečci oplodňují samičky (Voroncov 1986). Dle Millera (1956 in Vysoký 1995) je jednoletá generace, ale během dvou let mohou být tři pokolení. Podobně jako u jiných druhů není dostatečná zásoba chámových buněk pro oplodnění všech nakladených vajíček. Dochází k opakované kopulaci se samečky, kteří často hynou krátce po páření. Pfeffer (1954 in Vysoký 1995) druhé páření vylučuje. K oplození dochází v matečných chodbách, kde se samička vylíhla. Generační poměry nejsou jasné, ale podobně jako u většiny ostatních drtníků, se vyskytuje velmi málo samečků. Samečci jsou v menším počtu, asi 1:25 (Vysoký 1995). Na jednoho samečka tak připadá zpravidla 20–30 samiček (Voroncov 1986). Kultury hub v mateřských a larvových chodbách brzy hnědnou i při žíru. Samička na konci chodeb vyklade 20–30 vajíček. Vylíhlé larvy žerou pohromadě a vykusují ve dřevě nepravidelnou dutinu o ploše 2–19 cm2 směrem nahoru i dolů od matečné chodby. V malých chodbách je 20–30 larev, ve velkých dutinách pozorujeme až 200 larev pohromadě, ale zde se na snůšce podílí více samic. Někdy po hromadě více vyhlodaných dutin (Vysoký 1995).
3.3.2.4. Vajíčko
Vajíčko měkké a velké 0,7–0,8 x 0,35 mm (Vysoký 1995).
3.3.2.5. Larva
Larva je velká kolem 2 mm (Kutina 1983, Vysoký 1995) a je podobná druhu Xyleborus dispar, ale při pohledu ze strany se jeví pronotum zploštělé (Vysoký 1995).
3.3.2.6. Kukla
Kukla volná (pupa libera). Kuklí se v běli (Kalina 1999, 2004).
3.3.2.7. Příznaky napadení
Shodné s Xyleborus dispar.
3.3.2.8. Popis požerku
Tvoří nejjednodušší požerky (Voroncov 1986). Vstupní chodby jsou o průměru kolem 1 mm (0,5 mm – Gusev et Rimskij-Korsakov 1953). Proniká hluboko do běli (Vysoký 1995). Kalina (2004) a Urban (2004) však uvádí pouze 1–3 cm hluboko do běli jdoucí chodbu. Na rozdíl od ostatních drtníků se jeho larvy živí zčásti i dřevem, neboť hlavně v měkkém jarním dřevě vyžírají ploché, kapsovité dutiny. Dřeviny osidluje často spolu s drtníkem ovocným a pozoruhodné hospodářské škody může působit i v ovocných školkách a sadech (Urban 2004).
Hlodá v běli téměř všech druhů listnáčů svislou dlaňovitě rozšířenou krátkou matečnou chodbu. Matečná chodba se buď větevnatě dělí v rovině kolmé k ose kmene, anebo se dlaňovitě rozšiřuje nahoru a dolů. Matečná chodba 1–3 mm dlouhá se dlaňovitě rozšiřuje nahoru a dolů. Drtinky při závrtu jsou práškovité (Kutina 1983, Pfeffer 1989). Matečná chodba má v průměru 0,8–0,9 mm (Pfeffer 1989). Larvové chodby jsou společné, rodinné (Gusev et Rimskij-Korsakov 1953). Jde o ploškovou rodinou chodbu navazující na vstupní chodbu (Kalina 2004). Dokončený požerek larev má tvar čočkovité plošky a je údajně velký až 20 cm2 (Voroncov 1986, Vysoký 1995).
3.3.2.9. Parazité
Ve srovnání s bělokazy je v případě drtníků známo podstatně méně jejich parazitoidů (Hluchý et al. 1997). Vysoký (1995) uvádí pouze jediného predátora Rhizophagus aeneus Richt. A to pouze v olši.
3.3.2.10. Rozšíření
Palaearkt, Kanárské ostrovy, severní Amerika. V ČR hojně od nížin do hor. Pouze v hraničních horských pásmech chybí (Roubal 1937 in Vysoký 1995). Schwenke 1974 ho uvádí v celé Evropě. Pfeffer (1989) uvádí, že jde o hojnější druh celého rodu.
3.4. Způsoby sledování a přirození nepřátelé kůrovcovitých
3.4.1. Cizopasníci a další nepřátelé
Velký význam pro přirozenou rovnováhu má skupina hmyzů žijících v nepřátelském poměru k hmyzu potenciálně škodlivému. Jsou to buď cizopasné, nebo dravé druhy. Cizopasný hmyz žije na úkor těl svých hostitelů, kdežto dravý hmyz usmrcuje jiné druhy, které pak požírá. Cizopasnictví je dvojího druhu – buď cizopasníci žijí na povrchu těla hostitelů, to je cizopasnictví vnější – ektoparazitismus, nebo žijí uvnitř těla hostitelů, to je cizopasnictví vnitřní – endoparazitismus, z vnitřních parazitů mají největší význam kuklice a lumci. Vnitřní cizopasníci se vyvíjejí jen v určitých vývojových stádiích hostitelských hmyzů. Jsou i druhy, které se vyvíjejí jen v určitém druhu hostitelů – cizopasníci specifičtí, nebo zase druhy cizopasníků, kteří napadají různé hostitele – cizopasníci polyfágní. Brzdou v rozmnožování cizopasníků jsou však opět cizopasníci, kteří žijí v nich samých, tzv. hyperparazitoidi (např. Horák 2007c). Například Kruger et Mills (1990 in Faccoli 2002) uvádí jako parazitoidy lýkožrouta smrkového (Ips tyhographus) Hymenoptera z čeledi Braconidae Coeloides bostrychorum Giraud, Dendrosoter middendorffi R. a z čeledi Pteromalidae Roptrocerus xylofagorum R. a Rhopalicus tutela Walker.
Význam hmyzožravých savců a ptáků je dobře znám. Je třeba zvláště podporovat hnízdění užitečného ptactva nejen vyvěšováním budek, ale především ponecháváním vhodných dutých stromů a vytvářením vhodnějších životních podmínek. Na náhlém zániku hmyzích kalamit se často podílejí cizopasné houby, které způsobují onemocnění hmyzu, tzv. mykózy, bakterie způsobující bakteriózy a viry způsobující virózy. Aby byl hmyz chráněn před nepřátelským okolím, buď se ukrývá, nebo jeho tělo je zbarveno tak, že splývá se svým okolím, popřípadě tvarem svého těla napodobuje větévku, pupen, ptačí trus, nebo barvou i tvarem připomíná druhy hmyzu vybaveného žahadlem apod. Tím je odváděna především pozornost hmyzožravého ptactva (Hašek et Křístek in Vyskot 1962).
Významnou skupinou uplatňující se v ochraně je hmyz dravý. Vysoký (1995) uvádí, že kůrovci mají řadu přirozených nepřátel. V první řadě jsou to ptáci (datlové, strakapoudi, žluny, brhlíci a jiné druhy) a častěji se jedná o jiné druhy hmyzu. Poletující kusy loví vážky, larvy pronásledují brouci čeledi Carabidae, Staphylinidae, Scaphidiidae, Nitidulidae, Cucujidae, Colydiidae, Histeridae, Pythidae, Cleridae. Z dalšího hmyzu jsou to např. dlouhošijky (Rhaphididae) a hlavně Hymenoptera. Dále jsou to některé ploštice (Heteroptera) nebo zástupci dvoukřídlého hmyzu (Diptera). K tomu je nutno připočíst různé mikroby, bakterie, cizopasné houby aj.
Voroncov (1986) uvádí, že k dravým druhům podkorního hmyzu patří představitelé mnohých čeledí brouků, např. pestrokrovečník mravenčí (Thanasimus formicarius L.), potemník (Corticeus fraxini L.), mušky kovolesklice z rodu Medetera, mnohé druhy drabčíků (Staphylinidae) aj. Kůrovci hostí i četné druhy parazitů z říše hmyzu, zejména představitele čeledí kovověnkovitých (Pteromalidae) a lumčíkovitých (Braconidae) z řádu blanokřídlých. I další autoři (Kalina 2004, Hůrka 2005) uvádí u některých brouků žijících pod kůrou dřevin, že jsou predátory sensu lato. Pokud uvádí konkrétní predovanou skupinu brouků, jsou to právě kůrovci. Tyto údaje jsou však často přebírány od jiných autorů a často se tak objevují názory neopřené o přímá pozorování. Jde o brouky, jejichž bionomie je známa jen povrchně a tak jsou dohady o jejich dravosti podloženy jen jejich výskytem, tvarem těla, vzhledem larev atd. U mnoha druhů není predace kůrovců zcela jednoznačná. Lze to odvodit z jejich bionomie, kdy se larvy sice vyskytují pod kůrou odumřelých nebo odumírajících listnatých stromů, často jsou v detritu pod kůrou již v době, kdy se zde již žádná vývojová stadia kůrovců nevyskytují.
U dalších druhů, které bývají uváděny jako predátoři (Hůrka 2005) jde o druhy vyznačující se tím, že jejich larvy nesou většinou podobně utvářené urogomfy (zadečkové přívěšky – štěty). Jde např. u čeledi Cucujidae o lesáka rumělkového (Cucujus cinnaberinus Scopoli), u čeledi Pythidae Pytho depressus L. U čeledi Rhizophagidae většina druhů údajně pronásleduje kůrovce, jedním z nich je druh Rhizophagus bipustulatus F. U čeledi Silvanidae je to velmi plochý brouk Uleiota planata L., dále Cryptolestes ferrugineus Stephens z čeledi Laemophloeidae, který je dokládán i z mouky. Z potemníků Tenebrionidae je to např. Corticeus fasciatus F., který žije především v chodbách červotočů. Z čeledi Colydiidae dřevožrout zejkovaný (Bitoma crenata F.). Dále je to z čeledi Pyrochroidae kromě Pyrochroa coccinea L. např. i Schizotus pectiniornis L. Dále mnoho zástupců čeledi Melyridae, pestrokrovečníci z podčeledi Clerinae. A konečně dravá imaga a larvy podčeledi Trogossitinae např. Nemozoma elongatum L. nebo Temnoscheila coerulea Olivier. Kromě zástupců uvedených od čeledi Melyridae dále jde u valné většiny druhů pravděpodobně o saproxylofágy, saprofágy, mycetofágy, komenzály kůrovců nebo predátory jiných podkorních členovců.
3.4.2. Feromony
Obecně lze feromony rozdělit dle využití do tří základních kategorií (El-Sayed 2006).
Nejčastěji se používá feromon v kombinaci s lapacím zařízením. Souhrnný název je feromonový lapač (dále jen lapač). V ČR se nejčastěji používá lapač typu Theyson (štěrbinový) nebo Chemika 1 (křížový). Eventuelně lze v sadech využít i otrávený lapák z metrových polen ovocného stromu, na které je nastříkán kontaktní insekticid a připevněn feromon. Je vhodné postavit tři polena do trojnožky a pod ně umístit pro kontrolu bílou podložku. Jde však o metodu neselektivní a hubí se tak veškerý hmyz, který na lapák dosedne.
Tabulka 2: Feromonů a semiochemikálie prokázaných u kůrovců na ovocných dřevinách (dle El-Sayed 2006)
druh
název látky
kategorie
objevitel (rok)
Scolytus mali
5S7S-conophthorin
atraktant
Zhang (2002)
Scolytus rugulosus
ethyl alcohol
atraktant
Holgado (2000)
5S7S-conophthorin
atraktant
Zhang (2002)
Xyleborus dispar
ethyl alcohol
atraktant
Montgomery (1983)
ethyl alcohol
atraktant, vnadidlo
Klimetzek (1986)
atraktant
Martikainen (2001)
Xyleborus saxeseni
ethyl alcohol
atraktant
Roling (1975)
ethyl alcohol
atraktant, vnadidlo
Klimetzek (1986)
alpha-pinene
atraktant, vnadidlo
Petrice (2005)
beta-pinene
atraktant, vnadidlo
Petrice (2005)
4. DISKUSE
4.1. Vliv kůrovcovitých na ovocné dřeviny
Jak z rešerše vyplývá, za nejdůležitější zástupce z podčeledi kůrovcovitých (Scolytinae) na ovocných dřevinách lze považovat čtyři druhy kůrovců.
Za agresivnější jsou považováni druhy rodu bělokaz (Scolytus) a to vzhledem k jejich trofismu. Oba druhy rodu Scolytus jsou oligofágy (tzn., že jsou výživou omezení) čeledi růžovité (Rosaceae Juss.). Škodlivější je pravděpodobně druh bělokaz švestkový (Scolytus mali), který má širší potravní niku, co se tloušťky dřeva týče. Také se častěji vyskytuje na ovocných dřevinách. Bělokaz ovocný (Scolytus rugulosus) vystačí s tenčím materiálem, který v čeledi Rosaceae představují i ovocnářsky nezajímavé druhy jako například trnka (Prunus spinosa L.). Rod Scolytus svým žírem dokáže rostlinu postupně ubíjet, dřevina tak často živoří, zatímco pod kůrou je již téměř úplně rozežrána. Pokud jsou přítomny oba druhy rodu Scolytus, tak je dřevina zpravidla napadena až do poslední větve. Pokud se přidají krasci a tesaříci, nenajdeme často na stromě nenapadené místo.
Za méně agresivní a druhově specifické lze označit druhy rodu drtník (Xyleborus). Ten díky své strategii relativně méně poškozuje dřevinu. Jednak nezasahuje příliš do jejich vodivých cest a za druhé je jeho požerek plošně výrazně menší než u floeofágů (lýkožravých). Dalším faktorem je i to, že jde o polyfágy (všežravé), tzn., že danou dřevinu nepotřebuje ani tak kvůli úživnosti a látkám v ní obsažených, ale hlavně pro polohu dřeviny, tzn. oslunění, stáří nebo poškození. V užším slova smyslu by se dalo říci, že jde o druhy škodící na tvrdých listnáčích.
Za více agresivní lze označit druh drtník ovocný (Xyleborus dispar), ten napadá tenčí materiál a není takovým polyfágem. Škodlivým se může stát hlavně u školkařského materiálu, protože do přeroubovaných stromů na vzrůstných podnožích Xyleborus dispar příliš často nejde, u zákrsků na podnožích typu M9 se vyskytuje častěji, stromy na těchto podnožích jsou podstatně oslabenější (V. Kalina, os. sdělení). U stromů na podnoži M9 to zřejmě souvisí s tokem asimilátů, který jde lýkem přes jakýsi filtr a tím je strom brzděn v růstu, což je sice důležité u intenzivních výsadeb, strom to však potencionálně oslabuje.
Druhý z drtníků je drtník všežravý (Xyleborus saxeseni), ten se vyznačuje extrémní polyfágií, což v praxi znamená, že často dává přednost dřevinám v okolních stromových porostech. Jeho žír se dá označit za méně škodlivý, protože jde o druh s úzkou vazbou na ambróziovou houbu a rodinný požerek se velmi málo rozrůstá.
Je třeba zmínit ještě dalšího zástupce čeledi a to lýkohuba (Polygraphus grandiclava Thomson), ten je však vzhledem k nízké četnosti málo významný. Jde o polyfága, který napadá z ovocných dřevin třešně a višně. Napadá však také jehličnany, na ovocných dřevinách se tak jedná pravděpodobně o jinou potravní rasu.
U rodu Scolytus jde o fyziologické floeofágní škůdce, kteří ke svému vývoji nepotřebují symbiotické ambróziové (sloužící k výživě) houby. Přesto se však některé druhy hub v jejich mykoflóře nachází. Tito kůrovci nemají mycetangia a houby přenáší pasivně ve vnitřnostech a v různých záhybech kutikuly. Nejznámější druhy těchto hub jsou rod Ophiostoma, rod Ceratocystis a dále hyfomycety (vláknité houby) rodu Geosmithia, které tvoří bílý porost v požercích kůrovců. Tyto houby mohou tvořit přídatný zdroj potravy larev a dospělců nebo oslabovat napadenou dřevinu. Houby hrají důležitou roli v narušení obranného systému dřevin a mohou pomáhat rozkládat látky chránící dřevo. Kratší život floeofágních kůrovců oproti xylomycetofágním (ambróziovým) je přikládán jejich výživě lýkem, které je díky terpenům mírně toxické.
U rodu Xyleborus jde o fyziologicko-technické a technické xylomycetofágní škůdce dřevin. Ambrózioví brouci nehynou po ukončení rozmnožování, jako je tomu u floeofágních kůrovců. Dále spásají houbová mycelia a zatím neznámým způsobem udržují monokulturu ambróziové houby. Když dospělci ambróziových kůrovců zahynou, larvy dál také žít nemohou, protože neumějí udržovat čistotu houbové zahrádky. Larvy a brouci se živí podhoubím ambróziových hub Ambrosiella hartigii Batra a rodu Fusarium, které pokrývají stěny matečných chodeb.
Oba rody přednostně napadají oslabené dřeviny a to buď abiotickými činiteli jako sucho a mráz nebo biotickými jako mechanické poškození, přesazení, zkrácení kořenů, žír hlodavců, nevhodné hnojení atd. Jde o sekundární škůdce, o čemž svědčí i jejich výskyt v čerstvě pokácených dřevinách a u drtníků i v pařezech. Za příznivých klimatických a trofických podmínek se můžou přemnožit a napadnout dřeviny zdravé. Napadené dřeviny poté napadají i jiní hmyzí škůdci jako krasci, tesaříci a patogenní houby (např. ohňovec ovocný Phellinus pomaceus (Pers.) Maire).
4.1.1. Shrnutí – pořadí druhů dle agresivity
1. Scolytus mali
2.–3. Scolytus rugulosus a Xyleborus dispar
4. Xyleborus saxeseni
4.2. Symptomy napadení
4.2.1. Nejdůležitější vnější příznaky
Nejnápadnějším příznakem napadení dřeviny jsou drtinky, které se objevují u závrtových otvorů. Již na základě počtu závrtových a výletových otvorů a drtinek lze za určitých podmínek rozlišit oba rody.
U rodu Scolytus je závrtových otvorů většinou více, jedná se o rod monogamní (jeden ♂+jedna ♀). Na jeden rodinný požerek připadá jeden vstupní a podle délky ramene požerku ještě několik větracích otvorů. Zpravidla tak jsou minimálně 2 otvory na požerek. Výletových otvorů je také více, zpravidla na každého vylétlého brouka připadá jeden. Odlišná je i barva drtinek. Drtinky jsou sice zpočátku světlejší, ale protože pocházejí z borky a lýka, tak velmi záhy tmavnou. Jejich množství je také menší, protože lýko je potravou rodu Scolytus.
U rodu Xyleborus je relativně méně závrtových otvorů. Pro jeden rodinný požerek je pouze jeden závrt, který je zároveň výletovým otvorem. Drtinky u otvoru jsou světlejší, protože pochází z dřevní části. Je jich také podstatně více, protože samičky čistí chodby během celého vývoje.
4.2.1.1. Shrnutí – nejdůležitější vnější symptomy jednotlivých rodů
Rod Scolytus – při náletu v V a VIII je relativně více závrtových otvorů, což závisí i na stupni obsazení dřeviny. Drtinky jsou hnědé a jsou hlavně na počátku žíru. V období líhnutí nových brouků v VIII a V je velké množství výletových otvorů.
Rod Xyleborus – v druhé polovině IV při náletu je relativně méně závrtových otvorů. Drtinky jsou žluté a za časové období jich je více, nenalezneme je pouze po deštích. V době líhnutí brouků na jaře otvory nepřibývají.
4.2.2. Další vnější příznaky
Vzhledem k tomu, že většinu kůrovců je velmi obtížné objevit vně dřeviny, je poměrně dobrým ukazatelem, že strom je obsazen rodem Scolytus přítomnost brouků na borce během dne. Pro tento rod je totiž typické, že kopulují na povrchu dřevin a navíc jsou brouci nejaktivnější za dne. Jedince rodu Xyleborus lze nalézt většinou pouze v požerku, případně na začátku rojení ve večerních hodinách na borce při náletu na dřeviny. Samečci rodu Xyleborus jsou bezkřídlí a lze je nalézt pouze ve stromě, ve kterém se vylíhli.
Mezi další příznaky lze označit i vadnutí větviček nad místy žíru, což však lze při zběžnějším pohledu zaměnit s jinými poškozeními, jako například abiotické vadnutí suchem nebo poškození krupobitím, dále houbovými chorobami jako je moniliová spála (Monilia laxa (Ehrenb.) Sacc.) nebo nektriové usychání větví (Nectria cinnabarina (Tode) Fr.) ev. bakteriemi jako je rakovinné odumírání větví (Pseudomonas syringae), bakteriová spála růžovitých (Erwinia amylovora (Burill) Winsl.) ad. Nejlepším rozlišením je doba, kdy k vadnutí dochází a samozřejmě i symptomy.
4.2.3. Nejdůležitější vnitřní příznaky
Pro každý rod, případně i druh kůrovce je typický požerek. Základní rozdíl je, že u rodu Scolytus se nachází pod kůrou a u rodu Xyleborus se nachází ve dřevě.
Požerek druhů Scolytus získáme oddělením borky od dřevní části v místě závrtového otvoru. Nejlepší metodou je odloupnutí borky velikostí odpovídající asi 2/3 průměru kmene stromu. Plochu lze ohraničit obříznutím nožem a poté pomocí šroubováku odchlípnout. Pokud je rod přítomen, je přítomen i pro druh charakteristický požerek. Jediným rizikem je, že pokud vzorek odebíráme v době počínajícího rojení, může být pod kůrou pouze snubní komůrka nebo v lepším případě rameno požerku. Kromě tloušťky napadeného materiálu lze rozlišit požerek druhu Scolytus mali od S. rugulosus, tak že u S. mali je rameno požerku rovnější a rovnoběžné s osou kmene, snubní komůrka je s rozšířením na začátku, dále je u S. mali podstatně více vajíček nebo larvových chodbiček u a to 30–60 larvových chodeb oproti 10–20 u S. rugulosus, u kterého jsou larvové chodby podstatně kratší a více klikaté.
Požerek druhů rodu Xyleborus zjistíme tím, že po oddělení borky nalezneme pouze závrtový otvor, který pokračuje dále do dřeva. Tvar požerku získáme nejlépe podélným a příčným rozříznutím ev. rozštípnutím dřeva v místě závrtu. Požerek tohoto rodu je uvnitř zbarven zpravidla tmavě, což způsobuje ambróziová houba, kterou se tito kůrovci živí. Požerek druhu Xyleborus saxeseni má čočkovitý tvar bez dalšího větvení. U druhu Xyleborus dispar vypadá požerek následovně – od vstupní chodby jsou matečné chodby prvního řádu, které probíhají na obě strany zhruba ve směru letokruhů, ve slabším materiálu často na jednu stranu. Od nich pak jsou v nepravidelných odstupech různě dlouhé a různě početné matečné chodby druhého řádu, které jsou alespoň v koncových úsecích rovnoběžné s dřevními vlákny. V tenkých kmíncích a větvích matečné chodby často vybíhají ihned z krátké vstupní chodby. Všechny chodby v požerku mají stejný průměr, protože je hlodá tatáž samička.
4.2.3.1. Shrnutí – nejdůležitější vnitřní symptomy
Rod Scolytus – požerek se nachází pod kůrou, rameno má víceméně rovnoběžný směr s osou kmene.
Rod Xyleborus – požerek se nachází ve dřevě a většinou je pokryt tmavým podhoubím ambroziové houby.
Pozor je třeba dát na determinaci u bělokazů. U kterých jsou kukelné komůrky v běli a mohou připomínat závrty drtníků. Je třeba například drátkem ověřit hloubku dírky. Pokud bude dírka mělká, jde o kukelnou kolébku bělokaza. Pokud je hluboká jde o požerek drtníka. Napovědět, že jde o bělkaze může to, že otvorů je nápadně mnoho a jsou pravidelně rozmístěné.
4.3. Stručný popis rodů
4.3.1. Scolytus
Dospělci bělokazů mají bříško ostře seříznuté ve směru od zadních nohou ke konci krovek, proto připomíná zadní konec těla dláto. Hlava je svrchu zpod štítu viditelná. Štít a krovky jsou plošší, což je dobře vidět z boku. Tykadla a nožky jsou červenavě hnědé. Druh Scolytus mali oproti Scolytus rugulosus je větší, lesklý a poněkud světlejší, štít má jen málo tečkovaný. Larvy jsou pod kůrou a kukly v kukelné komůrce zapuštěné do běle.
4.3.2. Xyleborus
Dospělci drtníků se od sebe mnohem více liší. Mají zpravidla na zadním konci těla prohlubeninu – kárku, někdy obklopenou zoubky a tvořící jakýsi trakářek nebo košíček. Ta však u druhu Xyleborus dispar chybí. Nápadný je pohlavní dimorfismus (samičky se od samečků liší). Štít a krovky jsou na konci zakulacené (kromě samečka Xyleborus dispar). Štít je vpředu vroubkovaný a hlava není zpod štítu vidět. Samečci jsou poměrně vzácní, bezkřídlí, světlejší a menší než samičky. U obou pohlaví jsou nožky a tykadla žlutavá. Larvy a kukly jsou v požercích v běli.
Cílem této rešerše bylo uceleně zpracovat a utřídit informace u opomíjených druhů kůrovcovitých z podčeledi Scolytinae Latreille troficky vázaných na ovocné dřeviny pěstované v ČR. Poněkud slabší je výsledek u druhu Xyleborus saxeseni Ratzeburg, kterému je v odborné literatuře věnována dle našeho názoru malá pozornost, což je vzhledem k jeho polyfágii a poměrně zajímavému způsobu žíru zvláštní. Vzhledem k tomu jak je druh rozšířený, chybí podrobnější poznatky i z jeho bionomie. Kromě tohoto druhu se na ovocných dřevinách živí ještě Xyleborus dispar Fabricius, Scolytus mali Bechstein a Scolytus rugulosus Müller. Jediný druh, který se živí na ovocných dřevinách a nebyl zpracován, je Polygraphus grandiclava Thomson. Jedním z důvodů je jeho poměrně vzácný výskyt, složitý trofismus a nedostatečné informace v odborné literatuře. Snahou bylo zároveň vytvořit rešerši, která by mohla sloužit pro další podrobnější zkoumání druhů. Na závěr je třeba ještě konstatovat, že ač jsou kůrovci na ovocných dřevinách poměrně opomíjeni mohou být za určitých podmínek pro život dřeviny totální.
7. PODĚKOVÁNÍ
Rádi bych na tomoto místě poděkovali V. Kalinovi, který nám poskytl mnohé rady, podporu a hůře dostupnou literaturu. Také bychom rádi poděkovali J. Vávrovi, B. Zbuzkovi a Z. Jindrovi za připomínky k textu, za čož bychom rádi poděkovali i M. Knížkovi, který nám pomohl i s literárním přehledem.
8. SEZNAM POUŽITÉ LITERATURY
BALACHOWSKY A .S. 1949: Faune de France 50. Coléoptères Scolytides. Paul Lechevalier, Paris.
Beaver R. A. 1989: Insect-fungus relationship in the bark and ambrosia beetles. In: Wilding, N., Collins, N. M., Hammond, P.M., Weber J.F. (eds.) Insect-fungus interactions. Academic Press, London.
Blažek J. ET AL. 1998: Ovocnictví. Květ, Praha.
Blažek J. 2001: Pěstujeme jabloně. Brázda, Praha.
DOMINIK J. ET STARZYK J. R. 2004: Owady uszkodzajace drewno. Panstwowe Wydawnictwo Rolnicze i Lesne, Warszawa.
El-Sayed A. M. 2006: The Pherobase – Database of Insect Pheromones and Semiochemicals. <http://www.pherobase.com>.
Gusev V. I. ET Rimskij-Korsakov M. N. 1953: Klíč k určování škůdců lesních a okrasných stromů a keřů evropské části SSSR. SZN, Praha.
Faccoli M. 2002: Winter mortality in sub-corticolous populations of Ips typhographus (Coleoptera, Scolytidae) and its parazitids in the south-eastern Alps. Journal of Pest Science 75:62-68.
FARRELL B. D., SEQUEIRA A. S., MEARA B. C. O., NORMARK B. B., CHUNG J. H. ET JORDAL B. H. 2001: The Evolution of Agriculture in Beetles (Curculionidae: Scolytinae and Platypodinae). Evolution 55: 2011-2027.
Hluchý M. ET AL. 1997: Obrazový atlas chorob a škůdců ovocných dřevin a révy vinné, ochrana ovocných dřevin a révy vinné v integrované produkci. Biocont Laboratory s. r. o., Brno.
Horák J. 2007a: Kůrovci mohou být nebezpeční i v ovocnictví. Zahradnictví 1:16-19.
Horák J. 2007b: Další druhy podkorního hmyzu poškozujícího ovocné dřeviny. Zahradnictví 4:20-21.
Horák J. 2007c: Užitečný hmyz v zahradnictví – nepřítel škodlivého podkorního hmyzu. Zahradnictví 7:36-37.
Horák J. 2007d: Chráněný a ohrožený hmyz ovocných sadů. Zahradnictví 8:32-33.
Hulcr J. 2004: Lýkožrout v lese pohledem entomologa. Lesnická práce 4:18-19.
Hůrka K. 2005: Brouci České a Slovenské republiky. Kabourek, Zlín.
Kalina V. 1999: Entomologie pro LI, skripta LF ČZU v Praze, Praha.
Kalina V. 2004: Ochrana lesů, skripta LF ČZU v Praze, Praha.
KARPIŇSKI J. J. ET STRAWIŇSKI K. 1948: Korniki ziem Polski. Annales Universitatis Mariae Curie-Sklodowska, Supplementum IV, Sectio C, Lublin,.
KOLK A. ET STARZYK J. R. 1996: Atlas szkodlivych owadow lesnych. Multico, Warszawa.
Kazda J. ET AL. 2003: Choroby a škůdci polních plodin, ovoce a zeleniny. Zemědělec, Praha.
KNÍŽEK M. 2008: Kůrovcovití, jejich taxonomie, rozšíření a hospodářský význam. In: Horák, J.(ed.) Brouci vázaní na dřeviny. Pardubický kraj a ČLS, Pardubice.
Kofránek V. 1950: Stručný přehled našich dřev, jejich charakteristické znaky, technické vlastnosti a upotřebení. SN, Praha.
Kolařík M. 2004: Fascinující svět podkorního hmyzu – houbové symbiózy. Živa 2:73–75.
Kudela M. 1970: Atlas lesního hmyzu, škůdci na jehličnanech. SZN, Praha.
Kutina J. ET AL. 1983: Encyklopedie pro zahrádkáře 1. SZN, Praha.
Lanák J., Šimko K. ET Vanek G. 1969: Atlas chorob a škůdců ovocných plodin, révy vinné a zeleniny. SZN, Praha.
Nienhaus F., Butin H. ET Böhmer B. 1998: Atlas chorob a škůdců okrasných dřevin. Brázda, Praha.
Novák V., Hrozinka F. ET Starý B. 1974: Atlas hmyzích škůdců lesních dřevin. SZN, Praha.
NUNBERG M. 1954: Klucze do oznaczania owadow Polski, Czesc XIX Chrzaszcze – Coleoptera, Zeszyt 99-100, Korniki – Scolytidae, Wyrynniki – Platypodidae. Panstwowe Wydawnictwo Naukowe, Warszawa.
PFEFFER A. 1942: Kůrovci žijící na listnáčích. Les 22: 1-19.
PFEFFER A. 1954: Lesnická Zoologie II. Státní zemědělské nakladatelství, Praha.
PFEFFER A. 1961: Ochrana lesů. Státní zemědělské nakladatelství, Praha.
PFEFFER A. 1995: Zentral- und westpaläarktische Borken- und Kernkäfer (Coleoptera: Scolytidae, Platypodidae). Pro Entomologia, Basel.
PFEFFER A. 1961: Ochrana lesů. Státní zemědělské nakladatelství, Praha.
Pfeffer A. 1989: Kůrovcovcovití Scolytidae a jádrohlodovití Platypodidae. Academia, Praha.
PFFEFER A. ET KNÍŽEK M. 1993: Scolytidae, pp. 153-158 In: JELÍNEK J. (ed.): Seznam československých brouků. Check-list of Czechoslovak Insects IV (Coleoptera). Folia Heyrovskyana, Suppl. 1: 3-172 (in Czech and English).
Rauleder H. 2003: Beobachtungen zum Flugverlauf des Kleinen und des Ungleichen Holzbohrers (Xyleborus saxeseni und dispar). Gesunde Pflanzen 55:53-61.
Rauleder H. 2004:Beobachtungen zum Flugverlauf des Ungleichen Holzbohrers und des Schwarzen Nutzholzborkenkäfers – Korrektur der Flugdaten des Ungleichen Holzbohrers. Gesunde Pflanzen 56:170-174.
Schwenke W. 1974: Die Forstschädlinge Europas II. Band. Käfer. Verlag Paul Parey, Hamburg und Berlin.
STARK V. N. 1952: Zhestkokrylye, Koroedy. Fauna SSSR. Akademia Nauk SSSR, Moskva, Leningrad.
Šrůtka P. 2006: Vztah kambiofágního a xylofágního hmyzu a jeho doprovodné mykobioty. Disertační práce, FLE ČZU v Praze, Praha.
Švestka M. ET AL. 1998: Praktické metody v ochraně lesa. Lesnická práce, Kostelec nad Černými lesy.
Urban J. 2004: Drtník ovocný (Xyleborus dispar) a jak se mu bránit. Zahradnictví 1:20-22.
Voroncov A. I. ET Červinková H. 1986: Škůdci dřeva. SZN, Praha.
Vyskot M. ET AL. 1962: Praktická rukověť lesnická I. díl. SZN, Praha.
Vysoký V. 1995: Přehled technicky škodícího hmyzu na dříví (stručný souhrn znalostí z dostupné literatury). Albis international, Ústí nad Labem.
WOOD S. L. 1982: The bark and ambrosia beetles of North and Central America (Coleoptera: Scolytidae), a taxonomic monograph. Great Basin Naturalist Memoirs (6). Brigham Young University, Provo, Utah.
Zahradník P. ET Knížek M. 2000: Lýkožrout smrkový (Ips typhographus L.). Příloha leták lesní ochranné služby, Lesnická práce 10:8.
Zahradník P. 2006: Oponentský posudek disertační práce Ing. Petra Šrůtky Vztah kambioxylofágního hmyzu a jeho doprovodné mykobioty. VÚLHM, Strnady.
Zhang Q., Tolasch T., Schlyter F. ET Francke W. 2002: Enantiospecific antennal response of bark beetles to spiroacetal (e)-conophthorin. Journal of Chemical Ecology 28:1839-1852.